
Occurrence of Fusarium spp. in Maize Grain Harvested in Portugal and Accumulation of Related Mycotoxins during Storage

 ,
,  ,
,  , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling of Maize Grain
2.2. Chemicals Reagents
2.3. Climatic Conditions
2.4. Monitoring of Barrels
2.5. Fungal Analyses
2.5.1. Sample Preparation
2.5.2. Fusarium spp. Morphological Identification
2.5.3. Fusarium spp. Molecular Identification
2.6. Quantification of Mycotoxins
2.6.1. Mycotoxin Extraction
2.6.2. Mycotoxin UHPLC-ToF-MS Analysis
2.6.3. Deoxynivalenol (DON) Analysis
2.7. Statistical Analyses
3. Results and Discussion
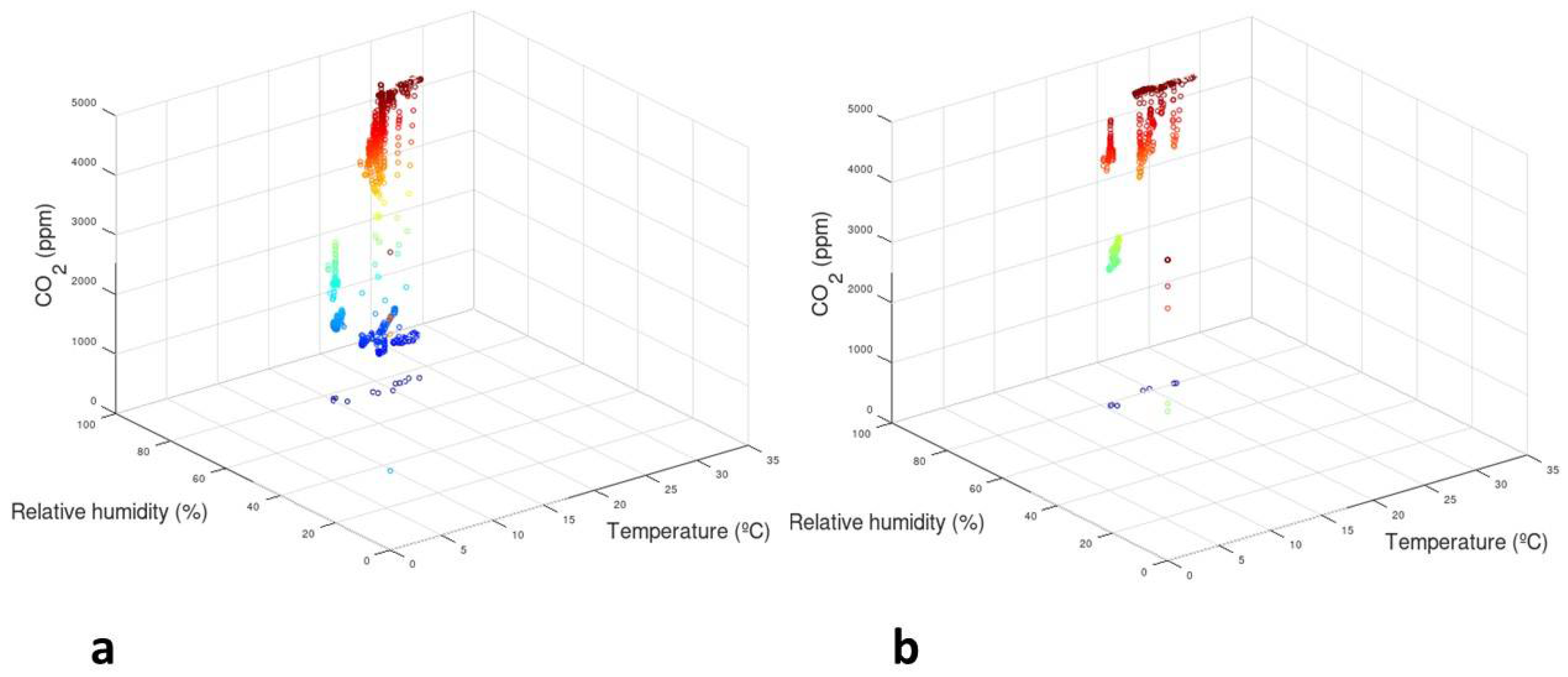
3.1. Monitoring of Maize Grain in Barrels
3.2. Fusarium spp. Presence in Maize
3.3. Mycotoxins Analyses
3.4. Correlations among Fumonisins, Fusarium Species, and Monitored Parameters
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Choi, J.H.; Lee, S.; Nah, J.Y.; Kim, H.K.; Paek, J.S.; Lee, S.; Ham, H.; Hong, S.K.; Yun, S.H.; Lee, T. Species composition of and fumonisin production by the Fusarium fujikuroi species complex isolated from Korean cereals. Int. J. Food Microbiol. 2018, 267, 62–69. [Google Scholar] [CrossRef]
- Dorn, B.; Forrer, H.R.; Jenny, E.; Wettstein, F.E.; Bucheli, T.D.; Vogelgsang, S. Fusarium species complex and mycotoxins in grain maize from maize hybrid trials and from grower’s fields. J. Appl. Microbiol. 2011, 111, 693–706. [Google Scholar] [CrossRef]
- INE -Instituto Nacional de Estatística, I.P. Estatísticas Agrícolas 2016; INE: Lisboa, Portugal, 2017; ISBN 9789892503950. [Google Scholar]
- INE- Instituto Nacional de Estatística, I.P. Estatísticas Agrícolas—2017; INE: Lisboa, Portugal, 2018; ISBN 978-989-25-0445-2. [Google Scholar]
- Eurostat Main Annual Crop Statistics. Eurostat statistics Explained. 2017. Available online: https://ec.europa.eu/eurostat/statistics-explained/index.php?title=Archive:Main_annual_crop_statistics&oldid=332968 (accessed on 19 December 2020).
- Pleadin, J.; Vahčić, N.; Perši, N.; Ševelj, D.; Markov, K.; Frece, J. Fusarium mycotoxins’ occurrence in cereals harvested from Croatian fields. Food Control. 2013, 32, 49–54. [Google Scholar] [CrossRef]
- Kirinčič, S.; Škrjanc, B.; Kos, N.; Kozolc, B.; Pirnat, N.; Tavčar-Kalcher, G. Mycotoxins in cereals and cereal products in Slovenia—Official control of foods in the years 2008–2012. Food Control. 2015, 50, 157–165. [Google Scholar] [CrossRef]
- Tima, H.; Brückner, A.; Mohácsi-Farkas, C.; Kiskó, G. Fusarium mycotoxins in cereals harvested from Hungarian fields. Food Addit. Contam. Part. B Surveill. 2016, 9, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Medina, Á.; Rodríguez, A.; Magan, N. Climate change and mycotoxigenic fungi: Impacts on mycotoxin production. Curr. Opin. Food Sci. 2015, 5, 99–104. [Google Scholar] [CrossRef]
- Gotthardt, M.; Asam, S.; Gunkel, K.; Moghaddam, A.F.; Baumann, E.; Kietz, R.; Rychlik, M. Quantitation of six Alternaria toxins in infant foods applying stable isotope labeled standards. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, B.; Forrer, H.R.; Schürch, S.; Vogelgsang, S. Fusarium species complex on maize in Switzerland: Occurrence, prevalence, impact and mycotoxins in commercial hybrids under natural infection. Eur. J. Plant. Pathol. 2009, 125, 51–61. [Google Scholar] [CrossRef]
- Boutigny, A.L.; Beukes, I.; Small, I.; Zühlke, S.; Spiteller, M.; Van Rensburg, B.J.; Flett, B.; Viljoen, A. Quantitative detection of Fusarium pathogens and their mycotoxins in South African maize. Plant. Pathol. 2012, 61, 522–531. [Google Scholar] [CrossRef]
- Ferrigo, D.; Raiola, A.; Causin, R. Fusarium toxins in cereals: Occurrence, legislation, factors promoting the appearance and their management. Molecules 2016, 21, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afsah-Hejri, L.; Jinap, S.; Hajeb, P.; Radu, S.; Shakibazadeh, S. A review on mycotoxins in food and feed: Malaysia case study. Compr. Rev. Food Sci. Food Saf. 2013, 12, 629–651. [Google Scholar] [CrossRef]
- Adekoya, I.; Obadina, A.; Phoku, J.; De Boevre, M.; De Saeger, S.; Njobeh, P. Fungal and mycotoxin contamination of fermented foods from selected south african markets. Food Control. 2018, 90, 295–303. [Google Scholar] [CrossRef]
- Hajok, I.; Kowalska, A.; Piekut, A.; Ćwieląg-Drabek, M. A risk assessment of dietary exposure to ochratoxin A for the Polish population. Food Chem. 2019, 284, 264–269. [Google Scholar] [CrossRef]
- Commission of the European Communities. Commission Regulation (EC) No 118/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 49, 5–24. [Google Scholar]
- Giorni, P.; Bertuzzi, T.; Battilani, P. Impact of fungi co-occurrence on mycotoxin contamination in maize during the growing season. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Vaughan, M.M.; Huffaker, A.; Schmelz, E.A.; Dafoe, N.J.; Christensen, S.; Sims, J.; Martins, V.F.; Swerbilow, J.; Romero, M.; Alborn, H.T.; et al. Effects of elevated [CO2] on maize defence against mycotoxigenic Fusarium verticillioides. Plant. Cell Environ. 2014, 37, 2691–2706. [Google Scholar] [CrossRef] [PubMed]
- Mshelia, L.P.; Selamat, J.; Samsudin, N.I.P.; Rafii, M.Y.; Abdul Mutalib, N.A.; Nordin, N.; Berthiller, F. Effect of temperature, water activity and carbon dioxide on fungal growth and mycotoxin production of acclimatised isolates of fusarium verticillioides and F. Graminearum. Toxins 2020, 12, 478. [Google Scholar] [CrossRef]
- Ezekiel, C.N.; Sulyok, M.; Ogara, I.M.; Abia, W.A.; Warth, B.; Šarkanj, B.; Turner, P.C.; Krska, R. Mycotoxins in uncooked and plate-ready household food from rural northern Nigeria. Food Chem. Toxicol. 2019, 128, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Zentai, A.; Szeitzné-Szabó, M.; Mihucz, G.; Szeli, N.; Szabó, A.; Kovács, M. Occurrence and risk assessment of fumonisin b1 and b2 mycotoxins in maize-based food products in Hungary. Toxins 2019, 11, 709. [Google Scholar] [CrossRef] [Green Version]
- Manizan, A.L.; Oplatowska-Stachowiak, M.; Piro-Metayer, I.; Campbell, K.; Koffi-Nevry, R.; Elliott, C.; Akaki, D.; Montet, D.; Brabet, C. Multi-mycotoxin determination in rice, maize and peanut products most consumed in Côte d’Ivoire by UHPLC-MS/MS. Food Control. 2018, 87, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Nie, D.; Fan, K.; Yang, J.; Guo, W.; Meng, J.; Zhao, Z.; Han, Z. A systematic review of plant-conjugated masked mycotoxins: Occurrence, toxicology, and metabolism. Crit. Rev. Food Sci. Nutr. 2020, 60, 1523–1537. [Google Scholar] [CrossRef] [PubMed]
- Tarazona, A.; Gómez, J.V.; Mateo, F.; Jiménez, M.; Romera, D.; Mateo, E.M. Study on mycotoxin contamination of maize kernels in Spain. Food Control. 2020, 118, 107370. [Google Scholar] [CrossRef]
- Kim, D.H.; Hong, S.Y.; Kang, J.W.; Cho, S.M.; Lee, K.R.; An, T.K.; Lee, C.; Chung, S.H. Simultaneous determination of multi-mycotoxins in cereal grains collected from South Korea by LC/MS/MS. Toxins 2017, 9, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kos, J.; Janić Hajnal, E.; Malachová, A.; Steiner, D.; Stranska, M.; Krska, R.; Poschmaier, B.; Sulyok, M. Mycotoxins in maize harvested in Republic of Serbia in the period 2012–2015. Part 1: Regulated mycotoxins and its derivatives. Food Chem. 2020, 312. [Google Scholar] [CrossRef] [PubMed]
- Kovač, M.; Bulaić, M.; Jakovljević, J.; Nevistić, A.; Rot, T.; Kovač, T. Mycotoxins, Pesticide Residues, and Heavy Metals Analysis of Croatian Cereals. Microorganisms 2021, 9, 216. [Google Scholar] [CrossRef]
- Abdallah, M.F.; Girgin, G.; Baydar, T.; Krska, R.; Sulyok, M. Occurrence of multiple mycotoxins and other fungal metabolites in animal feed and maize samples from Egypt using LC-MS/MS. J. Sci. Food Agric. 2017, 97, 4419–4428. [Google Scholar] [CrossRef]
- Getachew, A.; Chala, A.; Hofgaard, I.S.; Brurberg, M.B.; Sulyok, M.; Tronsmo, A.M. Multimycotoxin and fungal analysis of maize grains from south and southwestern Ethiopia. Food Addit. Contam. Part B Surveill. 2018, 11, 64–74. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, X.; Chen, G.; Sun, S.; Yang, Y.; Zhu, Z.; Duan, C. The major fusarium species causing maize ear and Kernel rot and their toxigenicity in Chongqing, China. Toxins 2018, 10, 90. [Google Scholar] [CrossRef] [Green Version]
- Mudili, V.; Siddaih, C.N.; Nagesh, M.; Garapati, P.; Naveen Kumar, K.; Murali, H.S.; Yli Mattila, T.; Batra, H.V. Mould incidence and mycotoxin contamination in freshly harvested maize kernels originated from India. J. Sci. Food Agric. 2014, 94, 2674–2683. [Google Scholar] [CrossRef]
- Oliveira, M.S.; Diel, A.C.L.; Rauber, R.H.; Fontoura, F.P.; Mallmann, A.; Dilkin, P.; Mallmann, C.A. Free and hidden fumonisins in Brazilian raw maize samples. Food Control. 2015, 53, 217–221. [Google Scholar] [CrossRef] [Green Version]
- Queiroz, V.A.V.; De Oliveira Alves, G.L.; Da Conceição, R.R.P.; Guimarães, L.J.M.; Mendes, S.M.; De Aquino Ribeiro, P.E.; Da Costa, R.V. Occurrence of fumonisins and zearalenone in maize stored in family farm in Minas Gerais, Brazil. Food Control. 2012, 28, 83–86. [Google Scholar] [CrossRef] [Green Version]
- García-Díaz, M.; Gil-Serna, J.; Vázquez, C.; Botia, M.N.; Patiño, B. A comprehensive study on the occurrence of mycotoxins and their producing fungi during the Maize production cycle in Spain. Microorganisms 2020, 8, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, A.S.; Brites, C.; Pouca, A.V.; Barbosa, J.; Freitas, A. UHPLC-ToF-MS method for determination of multi-mycotoxins in maize: Development and validation. Curr. Res. Food Sci. 2019, 1, 1–7. [Google Scholar] [CrossRef]
- Lino, C.M.; Silva, L.J.G.; Pena, A.L.S.; Silveira, M.I. Determination of fumonisins B1 and B2 in Portuguese maize and maize-based samples by HPLC with fluorescence detection. Anal. Bioanal. Chem. 2006, 384, 1214–1220. [Google Scholar] [CrossRef] [Green Version]
- Castellá, G.; Bragulat, M.R.; Rubiales, M.V.; Cabañes, F.J. Malachite green agar, a new selective medium for Fusarium spp. Mycopathologia 1997, 137, 173–178. [Google Scholar] [CrossRef]
- Thompson, R.S.; Aveling, T.A.S.; Blanco Prieto, R. A new semi-selective medium for Fusarium graminearum, F. proliferatum, F. subglutinans and F. verticillioides in maize seed. South. African J. Bot. 2013, 84, 94–101. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual Blackwell; Wiley-Blackwell: Oxford, UK, 2006; ISBN 9780813819198. [Google Scholar]
- Walsh, P.S.; Metzger, D.A.; Higuchi, R. Biotechniques 30th anniversary gem Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 2013, 54, 506–513. [Google Scholar] [CrossRef] [Green Version]
- Brewer, M.T.; Milgroom, M.G. Phylogeography and population structure of the grape powdery mildew fungus, Erysiphe necator, from diverse Vitis species. BMC Evol. Biol. 2010, 10, 268. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999. [Google Scholar] [CrossRef]
- Freitas, A.; Barros, S.; Brites, C.; Barbosa, J.; Silva, A.S. Validation of a Biochip Chemiluminescent Immunoassay for Multi-Mycotoxins Screening in Maize (Zea mays L.). Food Anal. Methods 2019, 12, 2675–2684. [Google Scholar] [CrossRef]
- Du, J.; Hu, M.; Zhang, W. Missing Data Problem in the Monitoring System: A Review. IEEE Sens. J. 2020, 20, 13984–13998. [Google Scholar] [CrossRef]
- Römer, D.; Bollazzi, M.; Roces, F. Carbon dioxide sensing in an obligate insectfungus symbiosis: CO2 preferences of leafcutting ants to rear their mutualistic fungus. PLoS ONE 2017, 12, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Magan, N.; Medina, A. Integrating gene expression, ecology and mycotoxin production by Fusarium and Aspergillus species in relation to interacting environmental factors. World Mycotoxin J. 2016, 9, 673–684. [Google Scholar] [CrossRef] [Green Version]
- Gromadzka, K.; Błaszczyk, L.; Chełkowski, J.; Waśkiewicz, A. Occurrence of mycotoxigenic fusarium species and competitive fungi on preharvest maize ear rot in poland. Toxins 2019, 11, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfordt, A.; Romero, L.R.; Schiwek, S.; Karlovsky, P.; von Tiedemann, A. Impact of environmental conditions and agronomic practices on the prevalence of fusarium species associated with ear-and stalk rot in maize. Pathogens 2020, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Ferrigo, D.; Raiola, A.; Causin, R. Plant stress and mycotoxin accumulation in maize. Agrochimica 2014, 58, 116–127. [Google Scholar] [CrossRef]
- Cao, A.; Santiago, R.; Ramos, A.J.; Souto, X.C.; Aguín, O.; Malvar, R.A.; Butrón, A. Critical environmental and genotypic factors for Fusarium verticillioides infection, fungal growth and fumonisin contamination in maize grown in northwestern Spain. Int. J. Food Microbiol. 2014, 177, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Balconi, C.; Berardo, N.; Locatelli, S.; Lanzanova, C.; Torri, A.; Redaelli, R. Evaluation of ear rot (Fusarium verticillioides) resistance and fumonisin accumulation in Italian maize inbred lines. Phytopathol. Mediterr. 2014, 53, 14–26. [Google Scholar] [CrossRef]
- Aguín, O.; Cao, A.; Pintos, C.; Santiago, R.; Mansilla, P.; Butrón, A. Occurrence of Fusarium species in maize kernels grown in northwestern Spain. Plant. Pathol. 2014, 63, 946–951. [Google Scholar] [CrossRef] [Green Version]
- Marín, S.; Magan, N.; Bellí, N.; Ramos, A.J.; Canela, R.; Sanchis, V. Two-dimensional profiles of fumonisin B1 production by Fusarium moniliforme and Fusarium proliferatum in relation to environmental factors and potential for modelling toxin formation in maize grain. Int. J. Food Microbiol. 1999, 51, 159–167. [Google Scholar] [CrossRef]
- Alberts, J.F.; Davids, I.; Moll, W.D.; Schatzmayr, G.; Burger, H.M.; Shephard, G.S.; Gelderblom, W.C.A. Enzymatic detoxification of the fumonisin mycotoxins during dry milling of maize. Food Control. 2020, 107726. [Google Scholar] [CrossRef]
- Vaughan, M.M.; Huffaker, A.; Schmelz, E.A.; Dafoe, N.J.; Christensen, S.A.; McAuslane, H.J.; Alborn, H.T.; Allen, L.H.; Teal, P.E.A. Interactive effects of elevated [CO2] and drought on the maize phytochemical defense response against mycotoxigenic Fusarium verticillioides. PLoS ONE 2016, 11, 1–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battilani, P.; Lanubile, A.; Scala, V.; Reverberi, M.; Gregori, R.; Falavigna, C.; Dall’asta, C.; Park, Y.S.; Bennett, J.; Borrego, E.J.; et al. Oxylipins from both pathogen and host antagonize jasmonic acid-mediated defence via the 9-lipoxygenase pathway in Fusarium verticillioides infection of maize. Mol. Plant. Pathol. 2018, 19, 2162–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandicke, J.; De Visschere, K.; Croubels, S.; De Saeger, S.; Audenaert, K.; Haesaert, G. Mycotoxins in flanders’ fields: Occurrence and correlations with fusarium species in whole-plant harvested maize. Microorganisms 2019, 7, 571. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sensor Variable | Range | Resolution | Precision |
|---|---|---|---|
| Relative humidity (%) | 0–100 | 0.01 | ±2 (0–90) |
| Temperature (°C) | −40–80 | 0.01 | 0.1@20 °C |
| CO2 (ppm) | 0–5000 | 1 | ±50 + 3% measured value |
| Plot | Storage Period | Relative Humidity (%) | Temperature (°C) | CO2 (ppm) |
|---|---|---|---|---|
| L2 | First 4 months | 71.2 | 15.2 | n.m. |
| Last 2 months | 68.5 | 20.1 | n.m. | |
| 6 Months | 71.3 (69.44–72.51) | 16.1 (13.77–21.95) | n.m. | |
| JC3 | First 4 months | 76.9 | 15.4 | 1554 |
| Last 2 months | 75.6 | 20.0 | 3816 | |
| 6 Months | 75.8 (72.87–78.93) | 19.3 (14.71–23.32) | 3437 (624–5000) | |
| JC4 | First 4 months | 77.8 | 15.6 | 3106 |
| Last 2 months | 80.3 | 20.5 | 4927 | |
| 6 Months | 79.9 (73.25–81.23) | 19.7 (14.85–24.11) | 4620 (2309–5000) |
| Fum B1 | Fum B2 | F. vert | F. subg | F. prolif | F. gram | F. spp. | Rel Hum | Temp | CO2 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Fum B1 | 0.96 ** | −0.75 * | 0.76 * | 0.66 | −0.16 | 0.17 | −0.54 | 0.45 | −0.26 | |
| Fum B2 | −0.67 * | 0.70 * | 0.60 | −0.16 | 0.14 | −0.65 | 0.22 | −0.43 | ||
| F. vert | −0.86 ** | −0.63 | −0.16 | −0.48 | 0.28 | −0.45 | 0.03 | |||
| F. subg | 0.43 | −0.32 | 0.59 | −0.22 | 0.51 | 0.03 | ||||
| F. prolif | 0.12 | −0.19 | −0.59 | 0.31 | −0.44 | |||||
| F. gram | −0.24 | 0.11 | −0.23 | 0.03 | ||||||
| F. spp. | 0.09 | 0.35 | 0.35 | |||||||
| Rel. Hum | 0.30 | 0.92 ** | ||||||||
| Temp | 0.61 | |||||||||
| CO2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carbas, B.; Simões, D.; Soares, A.; Freitas, A.; Ferreira, B.; Carvalho, A.R.F.; Silva, A.S.; Pinto, T.; Diogo, E.; Andrade, E.; et al. Occurrence of Fusarium spp. in Maize Grain Harvested in Portugal and Accumulation of Related Mycotoxins during Storage. Foods 2021, 10, 375. https://doi.org/10.3390/foods10020375
Carbas B, Simões D, Soares A, Freitas A, Ferreira B, Carvalho ARF, Silva AS, Pinto T, Diogo E, Andrade E, et al. Occurrence of Fusarium spp. in Maize Grain Harvested in Portugal and Accumulation of Related Mycotoxins during Storage. Foods. 2021; 10(2):375. https://doi.org/10.3390/foods10020375
Chicago/Turabian StyleCarbas, Bruna, Daniela Simões, Andreia Soares, Andreia Freitas, Bruno Ferreira, Alexandre R. F. Carvalho, Ana Sanches Silva, Tiago Pinto, Eugénio Diogo, Eugénia Andrade, and et al. 2021. "Occurrence of Fusarium spp. in Maize Grain Harvested in Portugal and Accumulation of Related Mycotoxins during Storage" Foods 10, no. 2: 375. https://doi.org/10.3390/foods10020375