Can Foliar Pulverization with CaCl2 and Ca(NO3)2 Trigger Ca Enrichment in Solanum tuberosum L. Tubers?

 ,
, 
 ,
,  , , , ,
, , , ,  , , , , , ,
, , , , , ,  , , ,
, , ,  and add
Show full author list
and add
Show full author list
Abstract
:1. Introduction
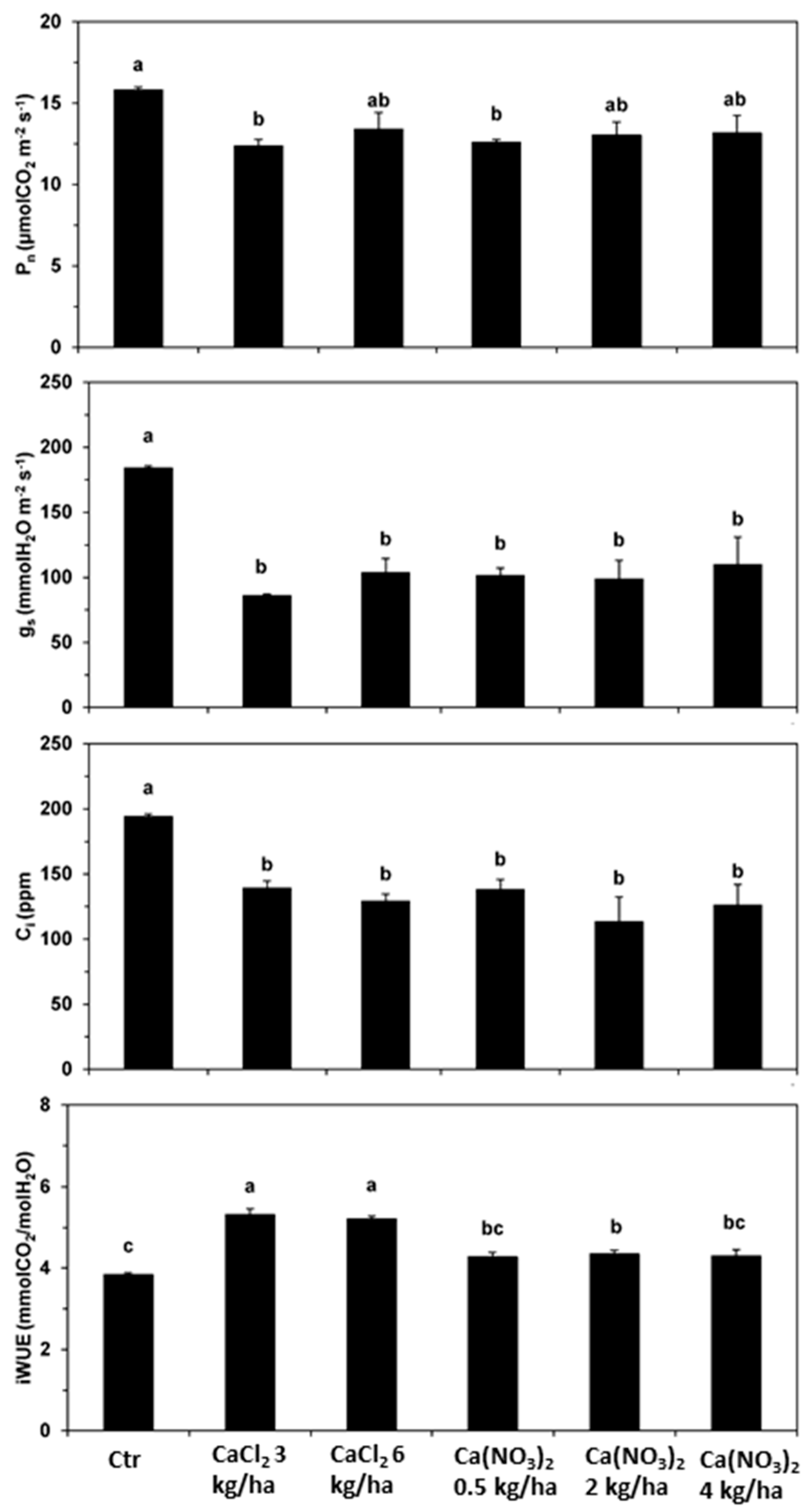
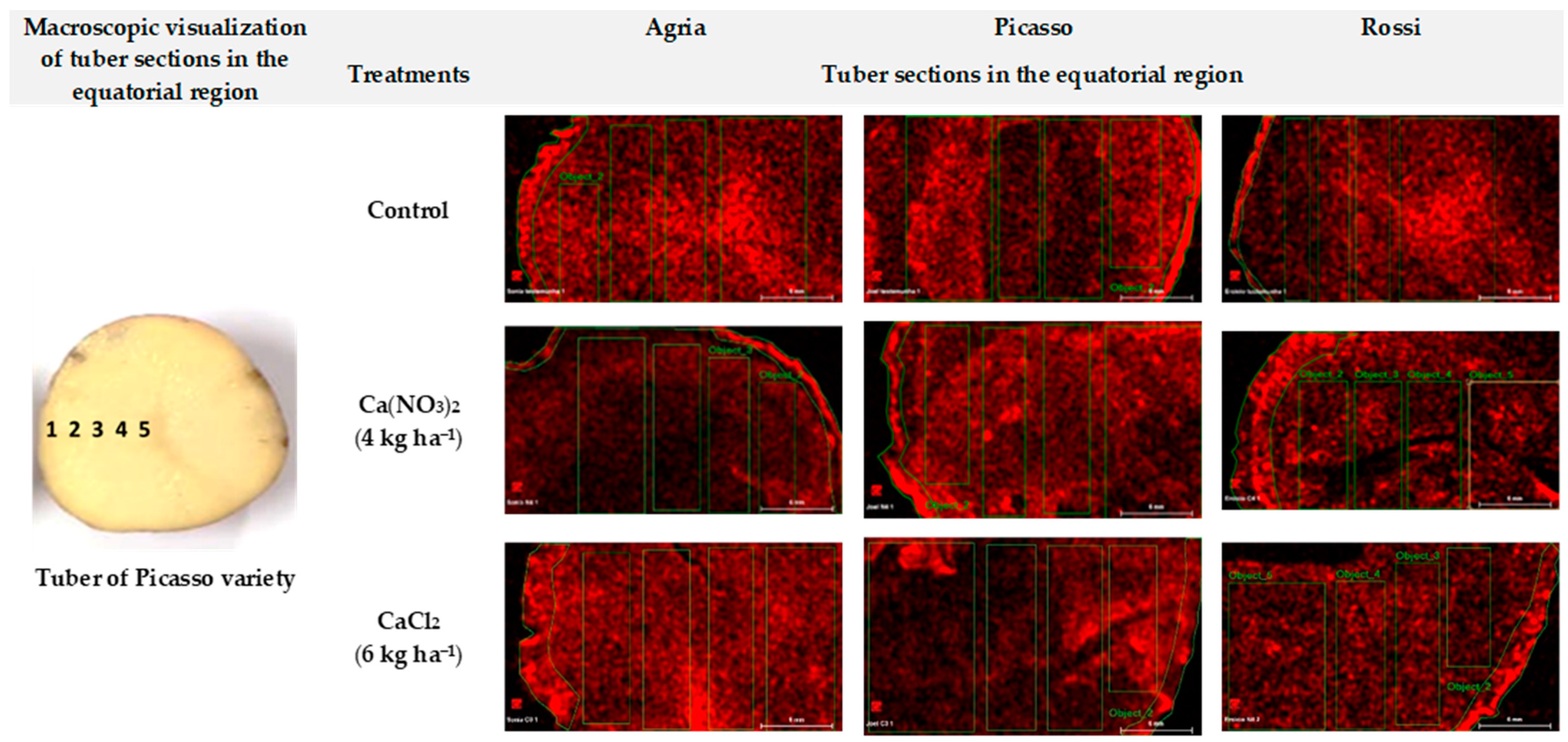
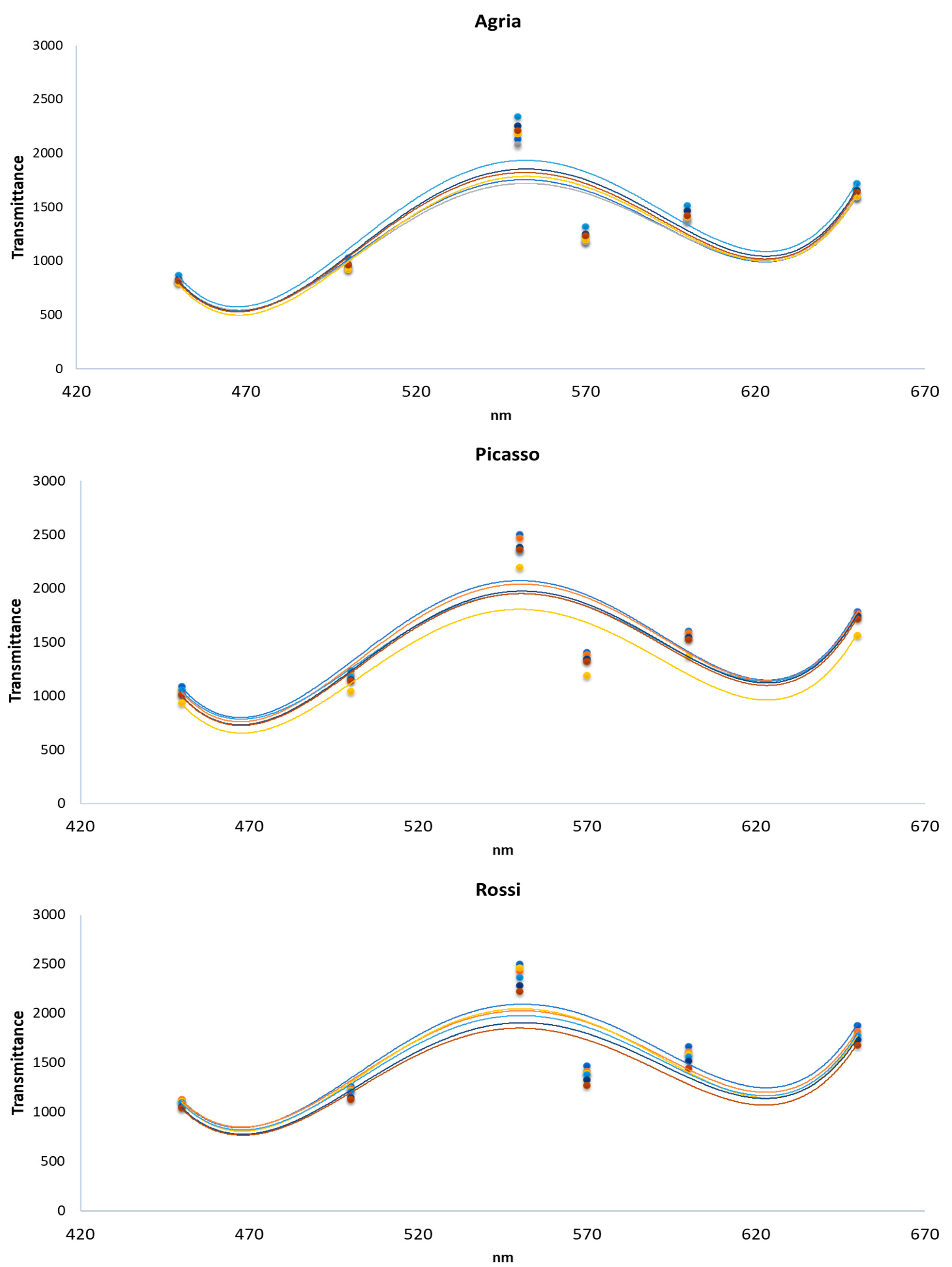
2. Results
3. Discussion
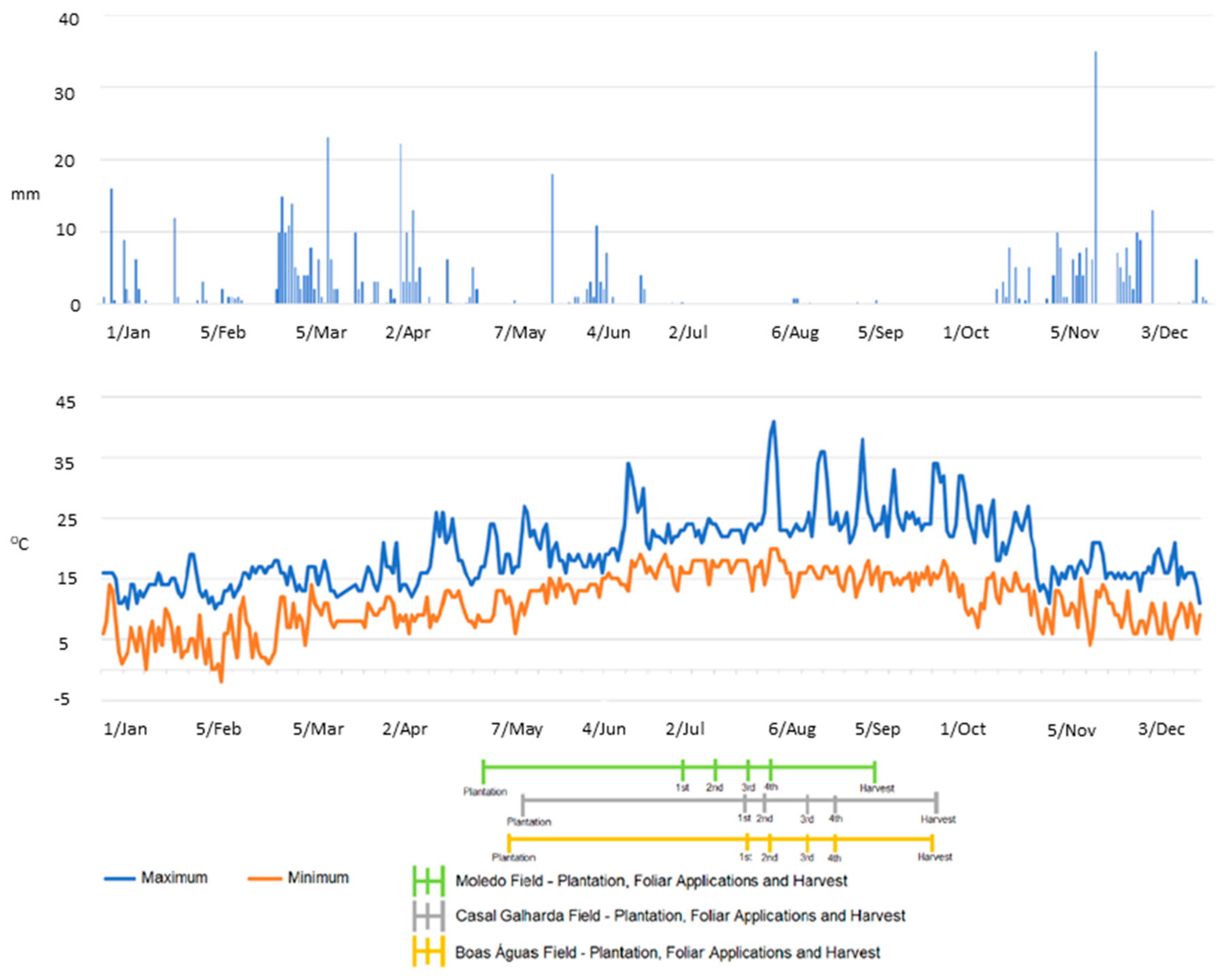
4. Materials and Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peacock, M. Calcium metabolism in health and disease. Clin. J. Am. Soc. Nephrol. 2010, 5 (Suppl. 1), S23–S30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IOM-Institute of Medicine. Dietary Reference Intakes for Calcium and Vitamin D; The National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-16395-8. [Google Scholar]
- Buchowski, M.S. Calcium in the context of dietary sources and metabolism. In Calcium: Chemistry, Analysis, Function and Effects; Preedy, V.R., Ed.; Food and Nutritional Components in Focus–Book Series; Royal Society of Chemistry: Cambridge, UK, 2015; Chapter 1; pp. 3–20. ISBN 978-1-78262-213-0. [Google Scholar] [CrossRef]
- Sharma, D.; Jamra, G.; Singh, U.; Sood, S.; Kumar, A. Calcium biofortification: Three pronged molecular approaches for dissecting complex trait of calcium nutrition in finger millet (Eleusine coracana) for devising strategies of enrichment of food crops. Front. Plant Sci. 2017, 7, 2028. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dayod, M.; Tyerman, S.; Leigh, R.; Gilliham, M. Calcium storage in plants and the implications for calcium biofortification. Protoplasma 2010, 247, 215–231. [Google Scholar] [CrossRef] [PubMed]
- EFSA (European Food Safety Authority). EFSA panel on dietetic products, nutrition and allergies. scientific opinion on dietary reference values for calcium. EFSA J. 2015, 13, 4101. [Google Scholar]
- Subramanian, N.K.; White, P.J.; Broadley, M.R.; Ramsay, G. The three-dimensional distribution of minerals in potato tubers. Ann. Bot. 2011, 107, 681–691. [Google Scholar] [CrossRef] [Green Version]
- Bouis, H.; Saltzman, A. Improving nutrition through biofortification: A review of evidence from HarvestPlus, 2003 through 2016. Glob. Food Segur. Agr. 2017, 12, 49–58. [Google Scholar] [CrossRef]
- García-Segovia, P.; Andrés-Bello, A.; Martínez-Monzó, J. Textural properties of potatoes (Solanum tuberosum L., cv. Monalisa) as affected by different cooking processes. J. Food Eng. 2008, 88, 28–35. [Google Scholar] [CrossRef]
- Lockyer, S.; White, A.; Buttriss, J. Biofortified crops for tackling micronutrient deficiencies-what impact are these having in developing countries and could they be of relevance within Europe? Nutr. Bull. 2018, 43, 319–357. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Roda, F.A.; Pessoa, M.F.G.; Reboredo, F.H.; Pais, I.P.; Ndayiragije, A.; Lidon, F.C.; Ribeiro-Barros, A.I. Selenium agronomic biofortification in rice: Improving crop quality against malnutrition. In The Future of Rice Demand: Quality Beyond Productivity; Costa de Oliveira, A., Pegoraro, C., Ebeling Viana, V., Eds.; Springer Nature: Cham, Switzerland, 2020; Chapter 8; pp. 179–203. ISBN 978-3-030-37509-6. [Google Scholar] [CrossRef]
- Oliveira, V.; Faquin, V.; Andrade, F.; Carneiro, J.; Júnior, E.; Souza, K.; Pereira, J.; Guilherme, L. Physiological and physicochemical responses of potato to selenium biofortification in tropical soil. Potato Res. 2019, 62, 315–331. [Google Scholar] [CrossRef]
- White, P.; Broadley, M. Biofortification of crops with seven mineral elements often lacking in human diets-iron, zinc, copper, calcium, magnesium, selenium and iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Wei, S.; Qin, G.; Zhang, H.; Tao, S.; Wu, J.; Wang, S.; Zhang, S. Calcium treatments promote the aroma volatiles emission of pear (Pyrus ussuriensis ‘Nanguoli’) fruit during post-harvest ripening process. Sci. Hortic. Amst. 2017, 215, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Hocking, B.; Tyerman, S.; Burton, R.; Gilliham, M. Fruit calcium: Transport and physiology. Front. Plant Sci. 2016, 7, 569. [Google Scholar] [CrossRef] [Green Version]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates, Inc.: Sunderland, UK, 2002; p. 665. [Google Scholar]
- White, P.J.; Broadley, M.R. Calcium in plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- White, P.J. The pathways of calcium movement to the xylem. J. Exp. Bot. 2001, 52, 891–899. [Google Scholar] [CrossRef]
- Moore, C.A.; Bowen, H.C.; Scrase-Field, S.; Knight, M.R.; White, P.J. The deposition of suberin lamellae determines the magnitude of sytosolic Ca2+ elevation in root endodermal cells subjected to cooling. Plant J. 2002, 30, 457–466. [Google Scholar] [CrossRef] [Green Version]
- Welch, R.M.; Shuman, L. Micronutrient nutrition of plants. CRC Crit. Rev. Plant Sci. 1995, 14, 49–82. [Google Scholar] [CrossRef]
- Barthakur, N.N.; Donnelly, D.J.; Habib, A. Transfer of strontium-90 and Ca-45 from medium to plant and their translocation in micropropagated potato. In Proceedings of the International Congress on the Radioecology and Ecotoxicology of Continental and Estuarine Environments (ECO-RAD, 2001), Aix en Provence, France, 3–7 September 2001. [Google Scholar]
- Atkinson, C.J. The Flux and Distribution of Xylem Sap Calcium to Adaxial and Abaxial Epidermal Tissue 8. J. Exp. Bot. 1991, 42, 987–993. [Google Scholar] [CrossRef]
- Atkinson, C.J.; Ruiz, L.P.; Mansfield, T.A. Calcium in xylem sap and the regulation of its delivery to the shoot. J. Exp. Bot. 1992, 43, 1315–1324. [Google Scholar] [CrossRef]
- Ziegler, H. Nature of transported substances. In Transport in Plants I: Phloem Transport. Encyclopedia of Plant Physiology; Zimmermann, M.H., Milburn, J.A., Eds.; Springer: New York, NY, USA, 1975; Volume 1, pp. 59–100. [Google Scholar]
- Welch, R.M. Importance of seed mineral nutrition reserves in crop growth and development. In Mineral Nutrition of Crops: Fundamental Mechanisms and Implications; Rengel, Z., Ed.; Food Products Press: New York, NY, USA, 1999; pp. 2005–2226. [Google Scholar]
- Ho, L.; White, P.J. A cellular hypothesis for the induction of blossom and rot in tomato fruit. Ann. Bot. 2005, 95, 571–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nelson, D.P.; Pan, W.L.; Franceschi, V.R. Xylem and phloem transport of mineral nutrients from Solanum tuberosum roots. J. Exp. Bot. 1990, 41, 1143–1148. [Google Scholar] [CrossRef]
- Davies, H.V.; Millard, P. Fractionation and distribution of calcium in sprouting and non-sprouting potato tubers. Ann. Bot. 1985, 56, 745–754. [Google Scholar] [CrossRef]
- Oparka, K.J.; Davies, H.V. Subcellular localisation of calcium in potato tubers. Potato Res. 1988, 31, 297–304. [Google Scholar] [CrossRef]
- Balamani, V.; Veluthambi, K.; Poovaiah, B.W. Effect of calcium on tuberization in potato (Solanum tuberosum L.). Plant Physiol. 1986, 80, 856–858. [Google Scholar] [CrossRef] [Green Version]
- Ewing, E.E. The Role of Hormones In Potato (Solanum tuberosum L.) Tuberization. In Plant Hormones; Davies, P.J., Ed.; Springer: Dordrecht, The Netherlands, 1995; pp. 698–721. [Google Scholar] [CrossRef]
- Jackson, S.D. Multiple signaling pathways control tuber induction in potato. Plant Physiol. 1999, 119, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ozgen, S.; Palta, J.P. Supplemental calcium application influences potato tuber number and size. HortScience 2004, 40, 102–105. [Google Scholar] [CrossRef] [Green Version]
- PEI-Nutrient Management, Prince Edward Island, Canada. 2014. Available online: www.gov.pe.ca/agriculture/labservices (accessed on 5 December 2020).
- El-Hadidi, E.M.; El-Dissoky, R.A.; Abdelhafez, A.A.H. Foliar calcium and magnesium application effect on potato crop grown in clay loam soils. JSSAE 2017, 8, 1–8. [Google Scholar] [CrossRef]
- El-Zohiri, S.S.M.; Asfour, H.E. Effects of foliar sprays of potassium, magnesium and calcium on yield, quality and storageability of potato. In Proceedings of the Fifth International Congress of Suastain Agriculture Development, Fayoum, Egypt, 21–23 December 2009; Faculty of Agriculture, Fayoum University: Fayoum, Egypt, 2009; pp. 57–71. [Google Scholar]
- Seifu, Y.W.; Deneke, S. Effect of calcium chloride and clacium nitrate on potato (Solanum tuberosum L.)growth and yield. J. Hortic. 2017, 4, 1–4. [Google Scholar]
- Teixeira, R.F.M.; Domingos, T.; Costa, A.P.S.V.; Oliveira, R.; Farropas, L.; Calouro, F.; Barradas, A.M.; Carneiro, J.P.B.G. Soil organic matter dynamics in Portuguese natural and sown rainfed grasslands. Ecol. Model. 2011, 222, 993–1001. [Google Scholar] [CrossRef]
- Koch, M.; Naumann, M.; Pawelzik, E.; Gransee, A.; Thiel, H. The importance of nutrient management for potato production Part I: Plant nutrition and yield. Potato Res. 2020, 63, 97–119. [Google Scholar] [CrossRef] [Green Version]
- D’Imperio, M.; Renna, M.; Cardinali, A.; Buttaro, D.; Serio, F.; Santamaria, P. Calcium biofortification and bioaccessibility in soilless “baby leaf” vegetable production. Food Chem. 2016, 213, 149–156. [Google Scholar] [CrossRef]
- Rowley, M.C.; Grand, S.; Verrecchia, E.P. Calcium-mediated stabilization of soil organic matter. Biogeochemistry 2018, 137, 27–49. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, R.; Miles, N.; Hughes, J.C. Interactions between potassium, calcium and magnesium in sugarcane grown on two contrasting soils in South Africa. Field Crops Res. 2018, 22, 1–11. [Google Scholar] [CrossRef]
- Kabata-Pendias, A. Trace Elements in Soils and Plants, 4th ed.; CRC Press: Boca Raton, FL, USA, 2010; 548p. [Google Scholar] [CrossRef]
- Klikocka, H.; Gáowacka, A. Does the Sulphur fertilization modify magnesium and calcium content in potato? Acta Sci. Pol. Hortorum Cult. 2013, 12, 41–53. [Google Scholar]
- Levy, D.; Fogelman, E.; Ytzak, Y. Influence of water and soil salinity on emergence and early development of potatoes (S. tuberosum L.) cultivar and effect of phisiological age of seed tubers. Potato Res. 1993, 36, 335–340. [Google Scholar] [CrossRef]
- Doorenbos, J.; Kassam, A.H. Efeito da Água No Rendimento das Culturas; UFPB: Campina Grande, Brazil, 1994; 306p. [Google Scholar]
- Levy, D. The response of potatoes (S. tuberosum L.) to salinity plant growth and tuber yields in the arid desert of Israel. Ann. Appl. Biol. 1992, 120, 547–555. [Google Scholar] [CrossRef]
- Ramalho, J.C.; Rebelo, M.C.; Santos, M.E.; Antunes, M.L.; Nunes, M.A. Effects of calcium deficiency on Coffea arabica. Nutrient changes and correlation of calcium levels with some photosynthetic parameters. Plant Soil 1995, 172, 87–96. [Google Scholar] [CrossRef]
- Hochmal, A.K.; Schulze, S.; Trompelt, K.; Hippler, M. Calcium-dependent regulation of photosynthesis. Biochim. Biophys. Acta 2015, 1847, 993–1003. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Yang, S.; Wan, S.; Li, X. The Significance of calcium in photosynthesis. Int. J. Mol. Sci. 2019, 20, 1353. [Google Scholar] [CrossRef] [Green Version]
- Allen, G.J.; Chu, S.P.; Harrington, C.L.; Schumacher, K.; Hoffmann, T.; Tang, Y.Y.; Grill, E.; Schroeder, J.I. A defined range of guard cell calcium oscillation parameters encodes stomatal movements. Nature 2001, 411, 1053–1057. [Google Scholar] [CrossRef]
- Weinl, S.; Held, K.; Schlücking, K.; Steinhorst, L.; Kuhlgert, S.; Hippler, M.; Kudla, J. A plastid protein crucial for Ca2+-regulated stomatal responses. New Phytol. 2008, 179, 675–686. [Google Scholar] [CrossRef]
- Drazeta, L.; Lang, A.; Hall, A.J.; Volz, R.K.; Jameson, P.E. Causes and effects of changes in xylem functionality in apple fruit. Ann. Bot. 2004, 93, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, H.; Fontes, P.; Finger, F.; Busato, C.; Cecon, P. Carboidratos e matéria seca de tubérculos de cultivares de batata influenciados por doses de nitrogênio. Cienc. Agrotec. 2010, 34, 285–293. [Google Scholar] [CrossRef]
- Virmond, E.; Kawakami, J.; Voncik, K.; Córdova, K.; Slompo, P. Características físico-químicas de cultivares de batata sob cultivo orgânico. Ambiência Guarapuava 2014, 10, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, A.; Soratto, R.; Evangelista, R.; Nardin, I. Qualidade físico-química e de fritura de tubérculos de cultivares de batata na safra de inverno. Hortic. Bras. 2010, 28, 299–304. [Google Scholar] [CrossRef] [Green Version]
- Direcção Geral de Agricultura Desenvolvimento Rural. Carta de Capacidade de uso do Solo de Portugal–Bases e Normas Adoptadas na sua Elaboração. Ministério da Economia, Secretaria de Estado da Agricultura, Serviço de Reconhecimento e de Ordenamento Agrário, 6ª Edição; República Portuguesa: Lisboa, Portugal, 1972; pp. 25–26. [Google Scholar]
- Pelica, J.; Barbosa, S.; Lidon, F.; Pessoa, M.F.; Reboredo, F.; Calvão, T. The paradigm of high concentration of metals of natural or anthropogenic origin in soils–the case of Neves-Corvo mine area (southern Portugal). J. Geochem. Explor. 2018, 186, 12–23. [Google Scholar] [CrossRef]
- Pessoa, F.P.; Scotti-Campos, P.; Pais, I.P.; Feiteiro, A.; Canuto, D.; Simões, M.; Pelica, J.; Pataco, I.; Ribeiro, V.; Reboredo, F.H.; et al. Nutritional profile of the Portuguese cabbage (Brassica oleracea L. var. Costata) and its relationship with the elemental soil analysis. Emir. J. Food Agric. 2016, 28, 381–388. [Google Scholar] [CrossRef] [Green Version]
- Rodier, J.; Legube, B.; Merlet, N. L’analyse de l’eau, 9th ed.; Dunod: Paris, France, 2009; p. 1579. ISBN 9782100072460. [Google Scholar]
- Piper, A.M. A graphic procedure in the geochemical interpretation of water analyses. Trans. Am. Geophys. Union. 1944, 25, 914–923. [Google Scholar] [CrossRef]
- Rodrigues, W.; Martins, M.; Fortunato, A.; Rodrigues, A.; Semedo, J.; Simões-Costa, M.; Pais, I.; Leitão, A.; Colwell, F.; Goulão, L.; et al. Long-term air [CO2] strenghtens photosymthetic functioning and mitigates the impact of supra-optimal temperatures in tropical Coffea arabica and C. canephora species. Glob. Chang. Biol. 2016, 22, 415–431. [Google Scholar] [CrossRef]
- Carrondo, M.; Reboredo, F.; Ganho, R.; Santos Oliveira, J.F. Heavy metal analysis of sediments in Tejo estuary, Portugal, using a rapid flameless atomic absorption procedure. Talanta 1984, 31, 561–564. [Google Scholar] [CrossRef]
- Reboredo, F.H.S.; Ribeiro, C.A.G. Vertical distribution of Al, Cu, Fe and Zn in soil salt marshes of the Sado estuary, Portugal. Int. J. Environ. Stud. 1984, 23, 249–253. [Google Scholar] [CrossRef]
- Cardoso, P.; Velu, G.; Singh, R.P.; Santos, J.P.; Carvalho, M.L.; Lourenço, V.M.; Lidon, F.C.; Reboredo, F.; Guerra, M. Localization and distribution of Zn and Fe in grains of biofortified bread wheat lines through micro- and triaxial- X-ray fluorescence spectrometry. Spectrochim. Acta B At. Spectrosc. 2018, 141, 70–79. [Google Scholar] [CrossRef]
- Mangueze, A.V.; Pessoa, M.F.G.; Silva, M.J.; Ndayiragije, A.; Magaia, H.E.; Cossa, V.S.I.; Reboredo, F.H.; Carvalho, M.L.; Santos, J.P.; Guerra, M.; et al. Simultaneous zinc and selenium biofortification in rice. Accumulation, localization and implications on the overall mineral content of the flour. J. Cereal Sci. 2018, 82, 34–41. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Field | Ability to Accumulate or Drain Surface Water | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Slope Class (%) | Surface Drainage | Area (m2) | Área (%) | ||||||||
| Moledo | 1-[0–5%] | Low | 238.0 | 53.7 | |||||||
| 2-[5–20%] | Moderate | 212.9 | 47.2 | ||||||||
| 3- >20% | High | 0.5 | 0.1 | ||||||||
| Casal Galharda | 1-[0–5%] | Low | 76.8 | 21.1 | |||||||
| 2-[5–20%] | Moderate | 265.5 | 72.9 | ||||||||
| 3- >20% | High | 21.7 | 6.0 | ||||||||
| Boas Águas | 1-[0–5%] | Low | 2.5 | 0.5 | |||||||
| 2-[5–20%] | Moderate | 475.4 | 99.5 | ||||||||
| 3- >20% | High | 0.1 | - | ||||||||
| Soil analysis (0–30 cm deep) (n = 16) | |||||||||||
| pH | Electrical Conductivity | Organic Matter | Ca | K | Mg | P | Fe | S | Zn | Mn | |
| μS cm −1 | % | ppm | |||||||||
| Moledo | 7.41 ± 0.03 a | 205 ± 17.5 b | 1.88 ± 0.12 c | 0.39 ± 0.03 b | 2.20 ± 0.03 b | 0.15 ± 0.01 b | 0.23 ± 0.01 b | 1.19 ± 0.07 b | 55.9 ± 4.80 b | 19.6 ± 1.54 c | 318 ± 26.5 b |
| Casal Galharda | 7.40 ± 0.05 a | 349 ± 24.2 a | 4.59 ± 0.10 a | 0.65 ± 0.03 a | 2.23 ± 0.07 b | 0.19 ± 0.02 b | 0.29 ± 0.01 a | 2.59 ± 0.06 a | 77.6 ± 1.11 a | 62.7 ± 2.77 a | 703 ± 56.8 a |
| Boas Águas | 7.30 ± 0.06 a | 332 ± 13.7 a | 4.13 ± 0.14 b | 0.71 ± 0.07 a | 2.64 ± 0.02 a | 0.24 ± 0.01 a | 0.19 ± 0.01 c | 0.50 ± 0.07 c | 66.6 ± 1.24 b | 41.7 ± 1.62 b | 270 ± 31.1 b |
| Water analysis | |||||||||||
| pH | Electrical Conductivity | Ca2± | K± | Mg2± | Na± | Cl− | HCO3− | SO42− | NO3− | PO43− | |
| μS cm −1 | mg L−1 (meq L−1) | ||||||||||
| Moledo | 7.2 | 1322 | 156.4(7.8) | 2.1 (0.05) | 22.5 (1.8) | 22.8 (1.4) | 56.6 (1.6) | 297.6 (4.8) | 166 (3.4) | 90.3 (1.4) | <1.5 (<0.04) |
| Casal Galharda | 6.9 | 1340 | 119.2(5.9) | 4.6 (0.1) | 37.1 (3.0) | 56.3 (2.4) | 89 (2.5) | 374.5 (6.1) | 164 (3.4) | 1.2 (0.01) | <1.5 (<0.04) |
| Boas Águas | 6.9 | 1381 | 169.6(8.4) | 2.2 (0.06) | 20.6 (1.7) | 41.2 (1.7) | 69 (1.9) | 330.6(5.4) | 234 (4.8) | 26.7 (0.4) | <1.5 (<0.04) |
| Treatments | Agria | Picasso | Rossi | |
|---|---|---|---|---|
| Ca (%) | ||||
| Control | 0.020 ± 0.001 b | 0.020 ± 0.001 c | 0.023 ± 0.000 a | |
| Ca(NO3)2 | 0.5 kg ha−1 | 0.021 ± 0.000 b | 0.021 ± 0.002 bc | 0.024 ± 0.001 ab |
| 2 kg ha−1 | 0.027 ± 0.000 a | 0.026 ± 0.001 ab | 0.025 ± 0.000 ab | |
| 4 kg ha−1 | 0.028 ± 0.001 a | 0.027 ± 0.001 ab | 0.026 ± 0.000 b | |
| CaCl2 | 3 kg ha−1 | 0.027 ± 0.002 a | 0.023 ± 0.001 abc | 0.023 ± 0.001 a |
| 6 kg ha−1 | 0.028 ± 0.000 a | 0.028 ± 0.002 a | 0.025 ± 0.001 ab | |
| Ca Contents (%) | |||||
|---|---|---|---|---|---|
| Treatments | Agria | ||||
| Transverse Sections in the Equatorial Region | |||||
| 1 | 2 | 3 | 4 | 5 | |
| Control | 0.31 ± 0.02 | 0.05 ± 0.00 | 0.07 ± 0.00 | 0.11 ± 0.01 | 0.09 ± 0.00 |
| Ca(NO3)2 (4 kg ha−1) | 0.82 ± 0.04 | 0.33 ± 0.02 | 0.26 ± 0.01 | 0.14 ± 0.01 | 0.16 ± 0.01 |
| CaCl2 (6 kg ha−1) | 0.46 ± 0.02 | 0.07 ± 0.00 | 0.11 ± 0.01 | 0.16 ± 0.01 | 0.36 ± 0.01 |
| Treatments | Picasso | ||||
| Transverse Sections in the Equatorial Region | |||||
| 1 | 2 | 3 | 4 | 5 | |
| Control | 1.03 ± 0.05 | 0.31 ± 0.02 | 0.07 ± 0.00 | 0.13 ± 0.01 | 0.47 ± 0.02 |
| Ca(NO3)2 (4 kg ha−1) | 1.02 ± 0.02 | 0.75 ± 0.04 | 1.02 ± 0.05 | 0.57 ± 0.03 | 0.66 ± 0.03 |
| CaCl2 (6 kg ha−1) | 1.00 ± 0.05 | 0.64 ± 0.03 | 0.34 ± 0.02 | 0.21 ± 0.01 | 0.33 ± 0.02 |
| Treatments | Rossi | ||||
| Transverse Sections in the Equatorial Region | |||||
| 1 | 2 | 3 | 4 | 5 | |
| Control | 0.34 ± 0.02 | 0.13 ± 0.01 | 0.17 ± 0.01 | 0.21 ± 0.01 | 0.15 ± 0.01 |
| Ca(NO3)2 (4 kg ha−1) | 0.90 ± 0.04 | 0.12 ± 0.01 | 0.13 ± 0.01 | 0.13 ± 0.01 | 0.22 ± 0.01 |
| CaCl2 (6 kg ha−1) | 0.15 ± 0.01 | 0.13 ± 0.01 | 0.11 ± 0.01 | 0.11 ± 0.01 | 0.24 ± 0.02 |
| Treatments | Agria | Picasso | Rossi | |
|---|---|---|---|---|
| Dry weight (%) | ||||
| Control | 25.2 ± 0.459 a | 20.9 ± 0.596 ab | 26.1 ± 1.180 a | |
| Ca(NO3)2 | 0.5 kg ha−1 | 21.0 ± 0.359 b | 17.6 ± 0.403 c | 18.1 ± 0.690 c |
| 2 kg ha−1 | 20.0 ± 0.248 b | 20.9 ± 0.533 ab | 21.6 ± 0.422 bc | |
| 4 kg ha−1 | 20.2 ± 0.660 b | 21.6 ± 0.403 ab | 22.8 ± 0.410 ab | |
| CaCl2 | 3 kg ha−1 | 22.6 ± 0.386 ab | 19.7 ± 0.158 bc | 23.2 ± 0.763 ab |
| 6 kg ha−1 | 22.4 ± 0.790 ab | 23.0 ± 0.713 a | 23.2 ± 0.090 ab | |
| Total Soluble Solids (°Brix) | ||||
| Control | 6.17 ± 0.136 a | 4.83 ± 0.136 b | 7.50 ± 0.408 a | |
| Ca(NO3)2 | 0.5 kg ha−1 | 5.33 ± 0.272 ab | 4.93 ± 0.054 b | 6.17 ± 0.136 bc |
| 2 kg ha−1 | 5.40 ± 0.249 ab | 5.17 ± 0.136 b | 5.17 ± 0.136 c | |
| 4 kg ha−1 | 5.00 ± 0.000 b | 5.50 ± 0.235 ab | 6.67 ± 0.134 ab | |
| CaCl2 | 3 kg ha−1 | 5.07 ± 0.054 ab | 6.51 ± 0.232 a | 7.33 ± 0.272 ab |
| 6 kg ha−1 | 5.30 ± 0.244 ab | 5.50 ± 0.288 ab | 7.00 ± 0.288 ab | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, A.R.F.; Lidon, F.C.; Pessoa, C.C.; Marques, A.C.; Luís, I.C.; Caleiro, J.; Simões, M.; Kullberg, J.; Legoinha, P.; Brito, M.; et al. Can Foliar Pulverization with CaCl2 and Ca(NO3)2 Trigger Ca Enrichment in Solanum tuberosum L. Tubers? Plants 2021, 10, 245. https://doi.org/10.3390/plants10020245
Coelho ARF, Lidon FC, Pessoa CC, Marques AC, Luís IC, Caleiro J, Simões M, Kullberg J, Legoinha P, Brito M, et al. Can Foliar Pulverization with CaCl2 and Ca(NO3)2 Trigger Ca Enrichment in Solanum tuberosum L. Tubers? Plants. 2021; 10(2):245. https://doi.org/10.3390/plants10020245
Chicago/Turabian StyleCoelho, Ana Rita F., Fernando C. Lidon, Cláudia Campos Pessoa, Ana Coelho Marques, Inês Carmo Luís, João Caleiro, Manuela Simões, José Kullberg, Paulo Legoinha, Maria Brito, and et al. 2021. "Can Foliar Pulverization with CaCl2 and Ca(NO3)2 Trigger Ca Enrichment in Solanum tuberosum L. Tubers?" Plants 10, no. 2: 245. https://doi.org/10.3390/plants10020245