
Comparative Genomics of Xanthomonas euroxanthea and Xanthomonas arboricola pv. juglandis Strains Isolated from a Single Walnut Host Tree

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Xanthomonas Strains Used in This Study and Genome Sequencing
2.2. Average Nucleotide Identity
2.3. Comparative Genome Analysis
2.4. Homologous of Pathogenicity and Virulence-Associated Proteins Inferred by tBLASTn Analysis
3. Results
3.1. General Features of X. euroxanthea and X. arboricola pv. juglandis Genome Assemblies
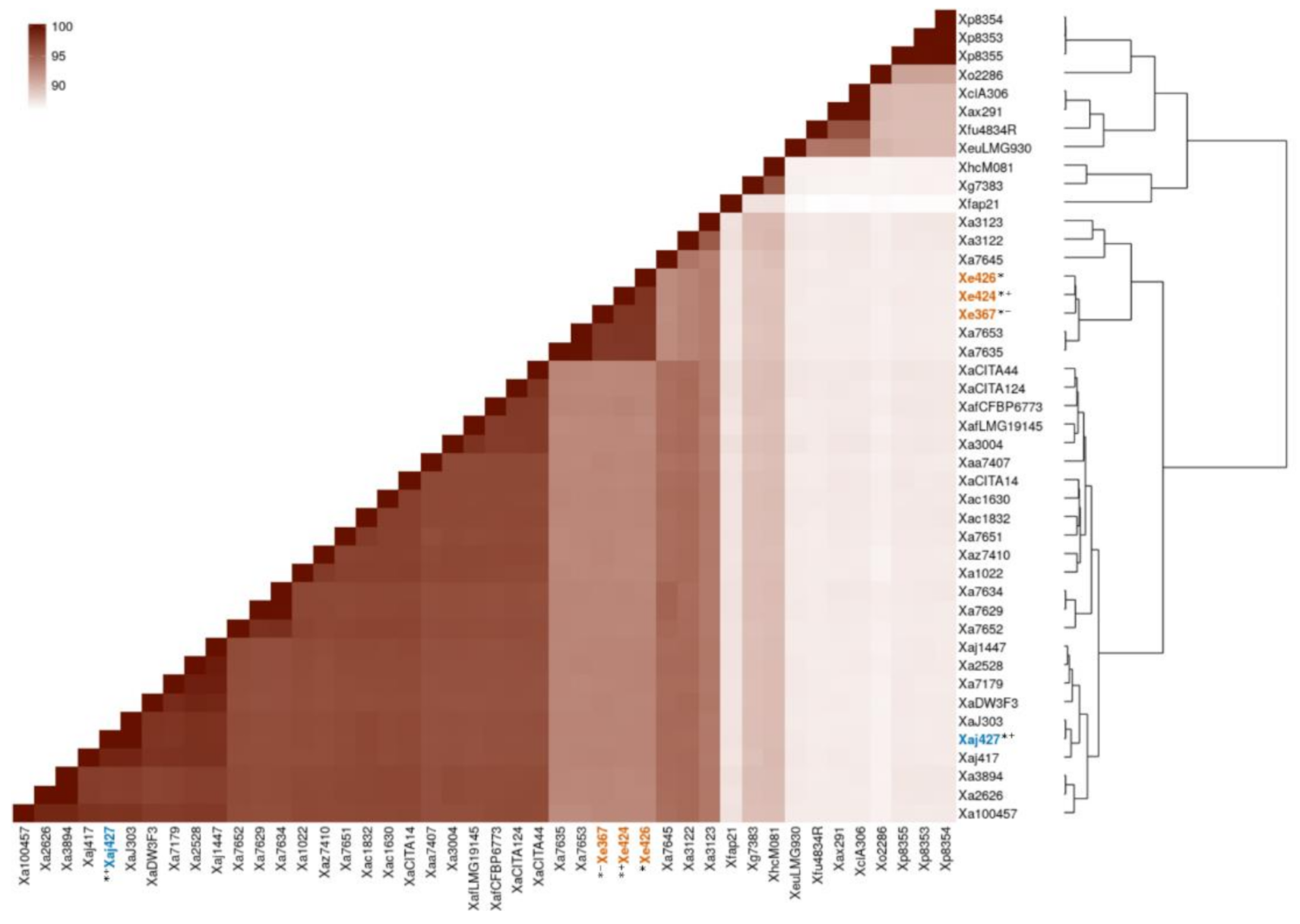
3.2. Genomic Distance Assessed by ANI and Phylogenetic Analysis
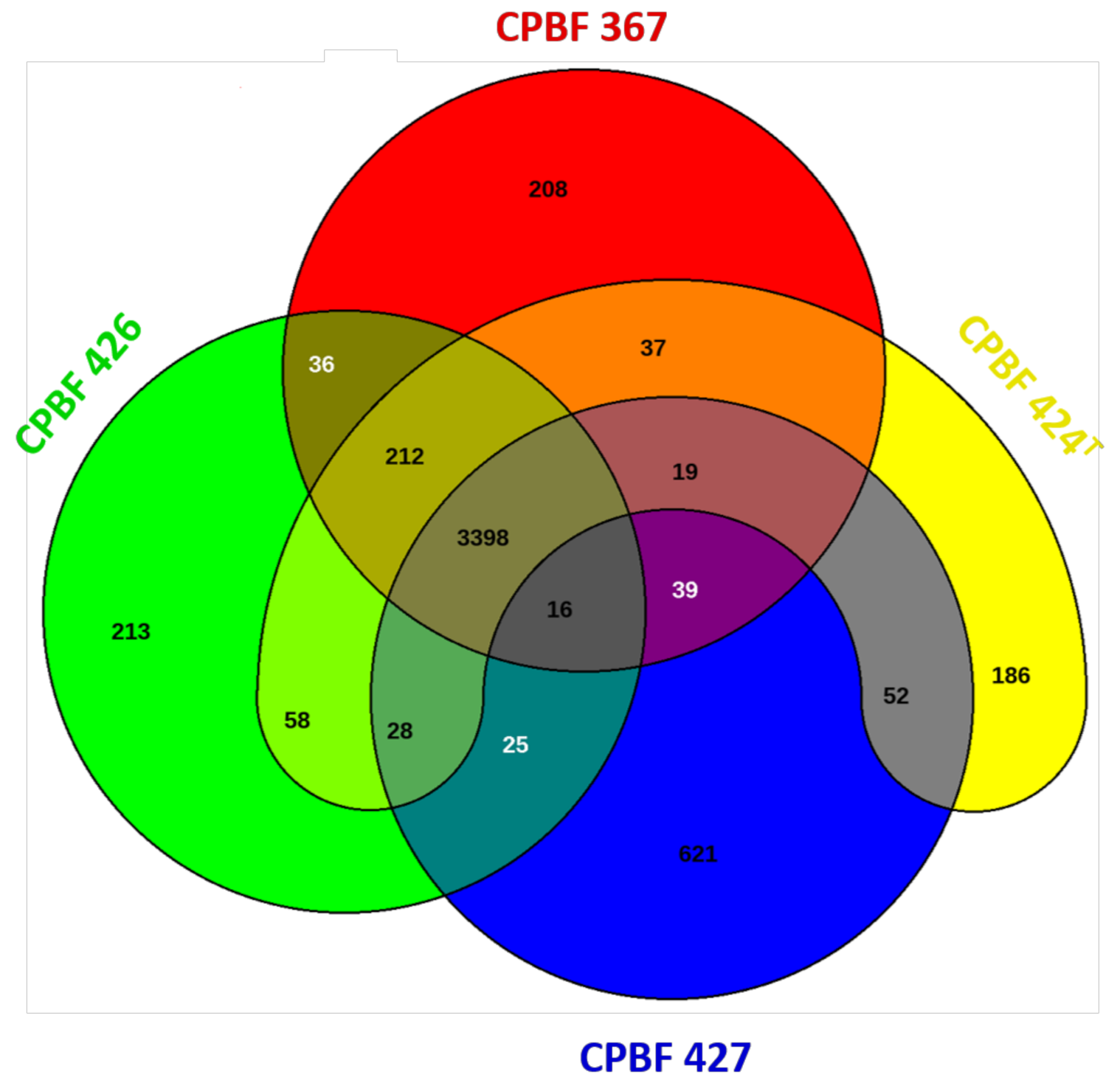
3.3. Genetic Patrimony Retrieved from the X. euroxanthea and X. arboricola pv. juglandis Strains Isolated from a Single Walnut Host Tree
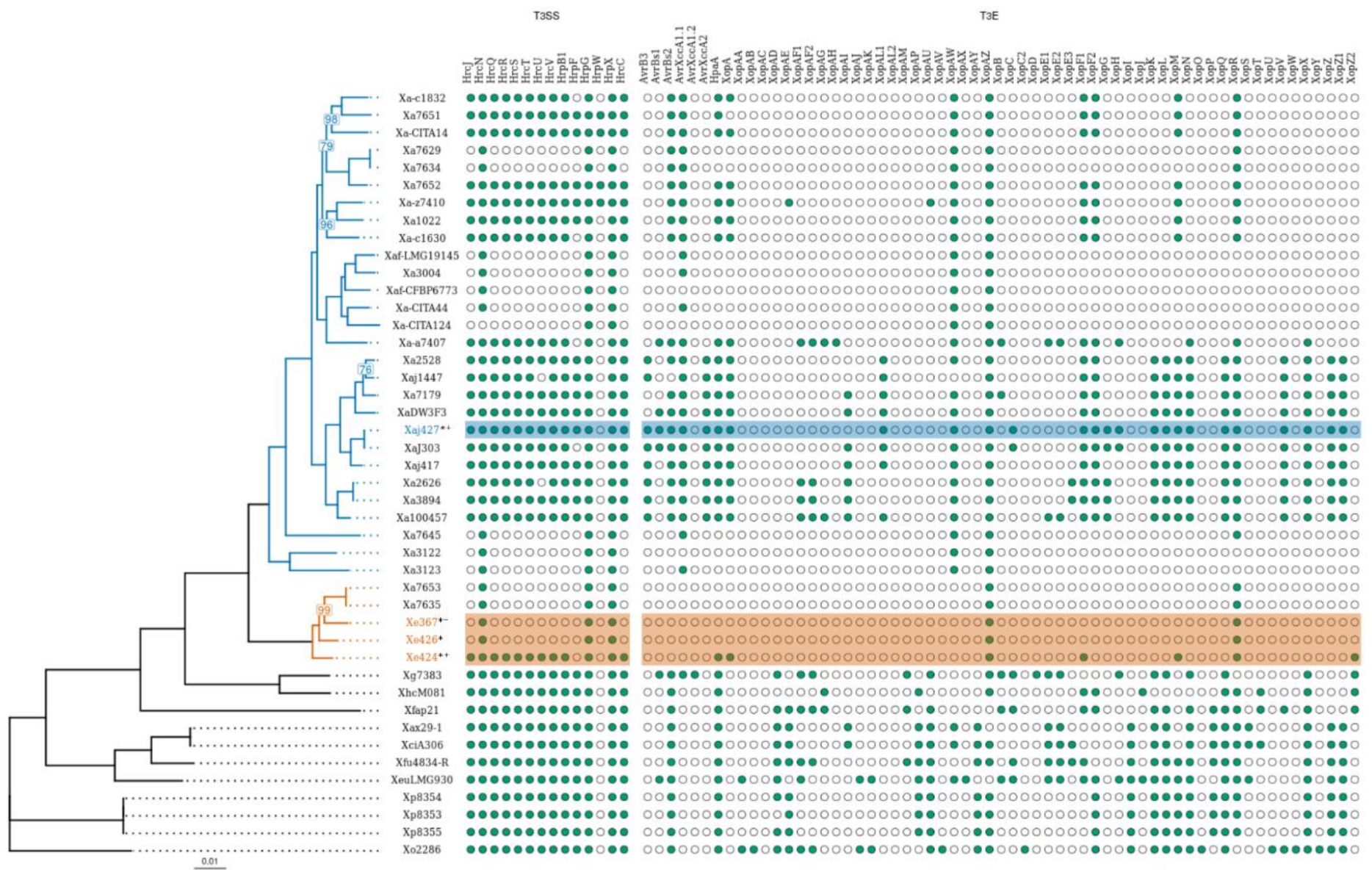
3.4. Pathogenic and Virulence-Related Factor Prediction
3.5. Type 3 Secretion System and Its Effectors
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dowson, D.W. On the systematic position and generic names of the Gram negative bacterial plant pathogens. Zentralblatt Fur Bakteriol. Parasitenkd. Und Infekt. 2 1939, 100, 177–193. [Google Scholar]
- Leyns, F.; De Cleene, M.; Swings, J.G.; De Ley, J. The host range of the genus Xanthomonas. Bot. Rev. 1984, 50, 308–356. [Google Scholar] [CrossRef]
- Swings, J.G.; Civerolo, E.L. Xanthomonas; Chapman & Hall: London, UK, 1993; ISBN 0412434202. [Google Scholar]
- Parte, A.C.; Carbasse, J.S.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic Names with Standing in Nomenclature (LPSN) Moves to the DSMZ. Available online: https://www.microbiologyresearch.org/content/journal/ijsem/10.1099/ijsem.0.004332 (accessed on 2 February 2021).
- Young, J.M.; Dye, D.W.; Bradbury, J.F.; Panagopoulos, C.G.; Robbs, C.F. A proposed nomenclature and classification for plant pathogenic bacteria. J. Agric. Res. 1978, 21, 153–177. [Google Scholar] [CrossRef]
- Vauterin, L.; Rademaker, J.; Swings, J. Synopsis on the taxonomy of the genus Xanthomonas. Phytopathology 2000, 90, 677–682. [Google Scholar] [CrossRef] [Green Version]
- Ryan, R.P.; Vorhölter, F.J.; Potnis, N.; Jones, J.B.; Van Sluys, M.A.; Bogdanove, A.J.; Dow, J.M. Pathogenomics of Xanthomonas: Understanding bacterium-plant interactions. Nat. Rev. Microbiol. 2011, 9, 344–355. [Google Scholar] [CrossRef]
- Meyer, D.F.; Bogdanove, A.J. Genomics-driven advances in Xanthomonas biology. In Genomics-Driven Advances in Xanthomonas Biology; Jackson, R., Ed.; Caister Academic Press: Norfolk, UK, 2009; pp. 147–161. [Google Scholar]
- Vauterin, L.; Hoste, B.; Kersters, K.; Swings, J. Reclassification of Xanthomonas. Int. J. Syst. Bacteriol. 1995, 45, 472–489. [Google Scholar] [CrossRef]
- Lamichhane, J.R. Xanthomonas arboricola diseases of stone fruit, almond, and walnut trees: Progress toward understanding and management. Plant Dis. 2014, 98, 1600–1610. [Google Scholar] [CrossRef] [Green Version]
- Pierce, N.B. Walnut bacteriosis. Bot. Gaz. 1901, 31, 272–273. [Google Scholar] [CrossRef]
- Smith, R.E.; Smith, C.O.; Ramsey, H.J. Walnut Culture in California: Walnut Blight; California Agricultural Experiments Station Publications: Berkeley, CA, USA, 1912. [Google Scholar]
- Hajri, A.; Meyer, D.; Delort, F.; Guillaumès, J.; Brin, C.; Manceau, C. Identification of a genetic lineage within Xanthomonas arboricola pv. juglandis as the causal agent of vertical oozing canker of Persian (English) walnut in France. Plant Pathol. 2010, 59, 1014–1022. [Google Scholar] [CrossRef]
- Belisario, A.; Maccaroni, M.; Corazza, L.; Balmas, V.; Valier, A. Occurrence and etiology of brown apical necrosis on Persian (English) walnut fruit. Plant Dis. 2002, 86, 599–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanović, Ž.; Popović, T.; Janse, J.; Kojić, M.; Stanković, S.; Gavrilović, V.; Fira, D. Molecular assessment of genetic diversity of Xanthomonas arboricola pv. juglandis strains from Serbia by various DNA fingerprinting techniques. Eur. J. Plant Pathol. 2014, 141, 133–145. [Google Scholar] [CrossRef]
- Kaluzna, M.; Pulawska, J.; Waleron, M.; Sobiczewski, P. The genetic characterization of Xanthomonas arboricola pv. juglandis, the causal agent of walnut blight in Poland. Plant Pathol. 2014, 63, 1404–1416. [Google Scholar] [CrossRef]
- Fischer-Le Saux, M.; Bonneau, S.; Essakhi, S.; Manceau, C.; Jacques, M.-A. Aggressive Emerging Pathovars of Xanthomonas arboricola Represent Widespread Epidemic Clones Distinct from Poorly Pathogenic Strains, as Revealed by Multilocus Sequence Typing. Appl. Environ. Microbiol. 2015, 81, 4651–4668. [Google Scholar] [CrossRef] [Green Version]
- Giovanardi, D.; Bonneau, S.; Gironde, S.; Fischer-Le Saux, M.; Manceau, C.; Stefani, E. Morphological and genotypic features of Xanthomonas arboricola pv. juglandis populations from walnut groves in Romagna region, Italy. Eur. J. Plant Pathol. 2016, 145, 1–16. [Google Scholar] [CrossRef]
- Fernandes, C.; Albuquerque, P.; Mariz-Ponte, N.; Cruz, L.; Tavares, F. Comprehensive diversity assessment of walnut-associated xanthomonads reveal the occurrence of distinct Xanthomonas arboricola lineages and of a new species (Xanthomonas euroxanthea) within the same tree. Plant Pathol. 2021, 1–16. [Google Scholar] [CrossRef]
- Essakhi, S.; Cesbron, S.; Fischer-Le Saux, M.; Bonneau, S.; Jacques, M.-A.; Manceau, C. Phylogenetic and Variable-Number Tandem-Repeat Analyses Identify Nonpathogenic Xanthomonas arboricola Lineages Lacking the Canonical Type III Secretion System. Appl. Environ. Microbiol. 2015, 81, 5395–5410. [Google Scholar] [CrossRef] [Green Version]
- Büttner, D.; Bonas, U. Regulation and secretion of Xanthomonas virulence factors. FEMS Microbiol. Rev. 2010, 34, 107–133. [Google Scholar] [CrossRef] [Green Version]
- An, S.Q.; Potnis, N.; Dow, M.; Vorhölter, F.J.; He, Y.Q.; Becker, A.; Teper, D.; Li, Y.; Wang, N.; Bleris, L.; et al. Mechanistic insights into host adaptation, virulence and epidemiology of the phytopathogen Xanthomonas. FEMS Microbiol. Rev. 2019, 44, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Hajri, A.; Pothier, J.F.; Le Saux, M.F.; Bonneau, S.; Poussier, S.; Boureau, T.; Duffy, B.; Manceau, C. Type three effector gene distribution and sequence analysis provide new insights into the pathogenicity of plant-pathogenic Xanthomonas arboricola. Appl. Environ. Microbiol. 2011, 78, 371–384. [Google Scholar] [CrossRef] [Green Version]
- Cesbron, S.; Briand, M.; Essakhi, S.; Gironde, S.; Boureau, T.; Manceau, C.; Fischer-Le Saux, M.; Jacques, M.-A. Comparative Genomics of Pathogenic and Nonpathogenic Strains of Xanthomonas arboricola Unveil Molecular and Evolutionary Events Linked to Pathoadaptation. Front. Plant Sci. 2015, 6, 1126. [Google Scholar] [CrossRef]
- Garita-Cambronero, J.; Palacio-Bielsa, A.; Cubero, J. Xanthomonas arboricola pv. pruni, causal agent of bacterial spot of stone fruits and almond: Its genomic and phenotypic characteristics in the X. arboricola species context. Mol. Plant Pathol. 2018, 19, 2053–2065. [Google Scholar] [CrossRef] [Green Version]
- Kchouk, M.; Gibrat, J.-F.; Elloumi, M. Generations of Sequencing Technologies: From First to Next Generation. Biol. Med. 2017, 9, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.; Giordano, F.; Ning, Z. Oxford Nanopore MinION Sequencing and Genome Assembly. Genom. Proteom. Bioinforma. 2016, 14, 265–279. [Google Scholar] [CrossRef] [Green Version]
- Oxford Nanopore Technologies Large Insights into Microorganisms. Available online: https://Nanoporetech.com/publication (accessed on 28 January 2021).
- Goldstein, S.; Beka, L.; Graf, J.; Klassen, J.L. Evaluation of strategies for the assembly of diverse bacterial genomes using MinION long-read sequencing. BMC Genom. 2019, 20, 23. [Google Scholar] [CrossRef] [Green Version]
- Martins, L.; Fernandes, C.; Blom, J.; Dia, N.C.; Pothier, J.F.; Tavares, F. Xanthomonas euroxanthea sp. nov., a new xanthomonad species including pathogenic and non-pathogenic strains of walnut. Int. J. Syst. Evol. Microbiol. 2020, 70, 6024–6031. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.; Blom, J.; Pothier, J.F.; Tavares, F. High-Quality Draft Genome Sequence of Xanthomonas sp. Strain CPBF 424, a Walnut-Pathogenic Strain with Atypical Features. Microbiol. Resour. Announc. 2018, 7. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.; Martins, L.; Fernandes, C.; Chaves, C.; Pinto, J.; Tavares, F.; Fonseca, N.A. Complete Genome Sequences of Walnut-Associated Xanthomonas euroxanthea Strains CPBF 367 and CPBF 426 Obtained by Illumina/Nanopore Hybrid Assembly. Microbiol. Resour. Announc. 2020, 9, e00902–20. [Google Scholar] [CrossRef]
- Fernandes, C.; Blom, J.; Pothier, J.F.; Tavares, F. High-Quality Draft Genome Sequence of Xanthomonas arboricola pv. juglandis CPBF 1521, Isolated from Leaves of a Symptomatic Walnut Tree in Portugal without a Past of Phytosanitary Treatment. Microbiol. Resour. Announc. 2018, 7, e00887–18. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.; Fernandes, C.; Chaves, C.; Pinto, J.; Tavares, F.; Fonseca, N.A. Complete Genome Sequence Obtained by Nanopore and Illumina Hybrid Assembly of Xanthomonas arboricola pv. juglandis CPBF 427, Isolated from Buds of a Walnut Tree. Microbiol. Resour. Announc. 2021, 10, e00085–21. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; Dicuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Lee, I.; Kim, Y.O.; Park, S.C.; Chun, J. OrthoANI: An improved algorithm and software for calculating average nucleotide identity. Int. J. Syst. Evol. Microbiol. 2016, 66, 1100–1103. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [Green Version]
- Waterhouse, R.M.; Seppey, M.; Simao, F.A.; Manni, M.; Ioannidis, P.; Klioutchnikov, G.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO applications from quality assessments to gene prediction and phylogenomics. Mol. Biol. Evol. 2018, 35, 543–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Edler, D.; Klein, J.; Antonelli, A.; Silvestro, D. RaxmlGUI 2.0: A graphical interface and toolkit for phylogenetic analyses using RAxML. Methods Ecol. Evol. 2021, 12, 373–377. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef]
- Yu, G.; Smith, D.K.; Zhu, H.; Guan, Y.; Lam, T.T. ggtree: An r package for visualization and annotation of phylogenetic trees with their covariates and other associated data. Methods Ecol. Evol. 2017, 8, 28–36. [Google Scholar] [CrossRef]
- Lerat, E.; Daubin, V.; Moran, N.A. From Gene Trees to Organismal Phylogeny in Prokaryotes:The Case of the γ-Proteobacteria. PLoS Biol. 2003, 1, e19. [Google Scholar] [CrossRef]
- Blom, J.; Albaum, S.P.; Doppmeier, D.; Pühler, A.; Vorhölter, F.J.; Zakrzewski, M.; Goesmann, A. EDGAR: A software framework for the comparative analysis of prokaryotic genomes. BMC Bioinform. 2009, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Blom, J.; Kreis, J.; Spänig, S.; Juhre, T.; Bertelli, C.; Ernst, C.; Goesmann, A. EDGAR 2.0: An enhanced software platform for comparative gene content analyses. Nucleic Acids Res. 2016, 44, W22–W28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Lee, B.M.; Park, Y.J.; Park, D.S.; Kang, H.W.; Kim, J.G.; Song, E.S.; Park, I.C.; Yoon, U.H.; Hahn, J.H.; Koo, B.S.; et al. The genome sequence of Xanthomonas oryzae pathovar oryzae KACC10331, the bacterial blight pathogen of rice. Nucleic Acids Res. 2005, 33, 577–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorhölter, F.J.; Schneiker, S.; Goesmann, A.; Krause, L.; Bekel, T.; Kaiser, O.; Linke, B.; Patschkowski, T.; Rückert, C.; Schmid, J.; et al. The genome of Xanthomonas campestris pv. campestris B100 and its use for the reconstruction of metabolic pathways involved in xanthan biosynthesis. J. Biotechnol. 2008, 134, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Darrasse, A.; Carrère, S.; Barbe, V.; Boureau, T.; Arrieta-Ortiz, M.L.; Bonneau, S.; Briand, M.; Brin, C.; Cociancich, S.; Durand, K.; et al. Genome sequence of Xanthomonas fuscans subsp. fuscans strain 4834-R reveals that flagellar motility is not a general feature of xanthomonads. BMC Genom. 2013, 14, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Crossman, L.; Dow, J.M. Biofilm formation and dispersal in Xanthomonas campestris. Microbes Infect. 2004, 6, 623–629. [Google Scholar] [CrossRef]
- Filloux, A. The underlying mechanisms of type II protein secretion. Biochim. Biophys. Acta Mol. Cell Res. 2004, 1694, 163–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczesny, R.; Jordan, M.; Schramm, C.; Schulz, S.; Cogez, V.; Bonas, U.; Büttner, D. Functional characterization of the Xcs and Xps type II secretion systems from the plant pathogenic bacterium Xanthomonas campestris pv. vesicatoria. New Phytol. 2010, 187, 983–1002. [Google Scholar] [CrossRef]
- Dunger, G.; Llontop, E.; Guzzo, C.R.; Farah, C.S. The Xanthomonas type IV pilus. Curr. Opin. Microbiol. 2016, 30, 88–97. [Google Scholar] [CrossRef]
- Shrivastava, S.; Mande, S.S. Identification and functional characterization of gene components of type VI secretion system in bacterial genomes. PLoS ONE 2008, 3, e2955. [Google Scholar] [CrossRef] [Green Version]
- Richter, M.; Rosselló-Móra, R. Shifting the genomic gold standard for the prokaryotic species definition. Proc. Natl. Acad. Sci. USA 2009, 106, 19126–19131. [Google Scholar] [CrossRef] [Green Version]
- Jacques, M.A.; Guimbaud, J.F.; Briand, M.; Indiana, A.; Darrasse, A. Flagellar Motility and Fitness in Xanthomonads. In Stress and Environmental Regulation of Gene Expression and Adaptation in Bacteria; Wiley Blackwell: Hoboken, NJ, USA, 2016; Volume 2, pp. 1265–1273. ISBN 9781119004813. [Google Scholar]
- Garita-Cambronero, J.; Palacio-Bielsa, A.; López, M.M.; Cubero, J. Comparative genomic and phenotypic characterization of pathogenic and non-pathogenic strains of Xanthomonas arboricola reveals insights into the infection process of bacterial spot disease of stone fruits. PLoS ONE 2016, 11, e0161977. [Google Scholar] [CrossRef] [PubMed]
- Jacques, M.-A.; Arlat, M.; Boulanger, A.; Boureau, T.; Carrère, S.; Cesbron, S.; Chen, N.W.G.; Cociancich, S.; Darrasse, A.; Denancé, N.; et al. Using Ecology, Physiology, and Genomics to Understand Host Specificity in Xanthomonas. Annu. Rev. Phytopathol. 2016, 54, 163–187. [Google Scholar] [CrossRef] [PubMed]
- Bansal, K.; Kumar, S.; Patil, P.B. Phylogenomic Insights into Diversity and Evolution of Nonpathogenic Xanthomonas Strains Associated with Citrus. mSphere 2020, 5, e00087-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quibod, I.L.; Perez-Quintero, A.; Booher, N.J.; Dossa, G.S.; Grande, G.; Szurek, B.; Vera Cruz, C.; Bogdanove, A.J.; Oliva, R. Effector Diversification Contributes to Xanthomonas oryzae pv. Oryzae Phenotypic Adaptation in a Semi-Isolated Environment. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef]
- Pereira, U.P.; Gouran, H.; Nascimento, R.; Adaskaveg, J.E.; Goulart, L.R.; Dandekar, A.M. Complete genome sequence of Xanthomonas arboricola pv. juglandis 417, a copper-resistant strain isolated from Juglans regia L. Genome. Genome Announc. 2015, 3, e01126-15. [Google Scholar] [CrossRef] [Green Version]
- Retamales, J.; Segovia, C.; Alvarado, R.; Nuñez, P.; Santander, J. Draft Genome Sequence of Xanthomonas arboricola pv. juglandis J303, Isolated from Infected Walnut Trees in Southern Chile. Genome Announc. 2017, 5, e01085–17. [Google Scholar] [CrossRef] [Green Version]
- Fu, B.; Chen, Q.; Wei, M.; Zhu, J.; Zou, L.; Li, G.; Wang, L. Complete Genome Sequence of Xanthomonas arboricola pv. juglandis Strain DW3F3, Isolated from a Juglans regia L. Bacterial Blighted Fruitlet. Genome Announc. 2018, 6, e00023–18. [Google Scholar] [CrossRef] [Green Version]
- Higuera, G.; González-Escalona, N.; Véliz, C.; Vera, F.; Romero, J. Draft Genome Sequences of Four Xanthomonas arboricola pv. juglandis Strains Associated with Walnut Blight in Chile: TABLE 1. Genome Announc. 2015, 3, e01160–15. [Google Scholar] [CrossRef] [Green Version]
- Garita-Cambronero, J.; Palacio-Bielsa, A.; López, M.M.; Cubero, J. Draft genome sequence for virulent and avirulent strains of Xanthomonas arboricola isolated from Prunus spp. in Spain. Stand. Genom. Sci. 2016, 11, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Ignatov, A.N.; Kyrova, E.I.; Vinogradova, S.V.; Kamionskaya, A.M.; Schaad, N.W.; Luster, D.G. Draft genome sequence of Xanthomonas arboricola strain 3004, a causal agent of bacterial disease on barley. Genome Announc. 2015, 3, 1572–1586. [Google Scholar] [CrossRef] [Green Version]
- Harrison, J.; Grant, M.R.; Studholme, D.J. Draft genome sequences of two strains of Xanthomonas arboricola pv. celebensis isolated from banana plants. Genome Announc. 2016, 4, e01705-15. [Google Scholar] [CrossRef] [Green Version]
- Medini, D.; Donati, C.; Tettelin, H.; Masignani, V.; Rappuoli, R. The microbial pan-genome. Curr. Opin. Genet. Dev. 2005, 15, 589–594. [Google Scholar] [CrossRef]
- Hersemann, L.; Wibberg, D.; Blom, J.; Goesmann, A.; Widmer, F.; Vorhölter, F.J.; Kölliker, R. Comparative genomics of host adaptive traits in Xanthomonas translucens pv. graminis. BMC Genom. 2017, 18, 35. [Google Scholar] [CrossRef] [Green Version]
- Garita-Cambronero, J.; Palacio-Bielsa, A.; López, M.M.; Cubero, J. Pan-genomic analysis permits differentiation of virulent and non-virulent strains of Xanthomonas arboricola that cohabit Prunus spp. and elucidate bacterial virulence factors. Front. Microbiol. 2017, 8, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Figueiredo, F.; Jones, J.; Wang, N. HrpG and HrpX Play Global Roles in Coordinating Different Virulence Traits of Xanthomonas axonopodis pv. citri. Mol. Plant Microbe Interact. 2011, 24, 649–661. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, J.M.; Pesce, C.; Lefeuvre, P.; Koebnik, R. Comparative genomics of a cannabis pathogen reveals insight into the evolution of pathogenicity in Xanthomonas. Front. Plant. Sci. 2015, 6, 431. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| General Features | Xanthomonas euroxanthea Strains | Xanthomonas arboricola pv. juglandis Strain | ||
|---|---|---|---|---|
| CPBF 367 | CPBF 424T | CPBF 426 | CPBF 427 | |
| Genome size (bp) | 4,968,459 | 4,900,930 | 4,900,648 | 5,228,174 |
| Contigs | 2 | 1 | 2 | 1 |
| N50 (bp) | 4,923,218 | 4,900,930 | 4,883,254 | 5,228,174 |
| G+C content (%) | 65.81 | 65.88 | 65.85 | 65.38 |
| Plasmids | 1 | 0 | 1 | 0 |
| Total genes | 4157 | 4119 | 4143 | 4465 |
| Total CDSs | 4077 | 4040 | 4066 | 4367 |
| Coding genes | 4012 | 3993 | 4003 | 4237 |
| RNA genes | 80 | 79 | 77 | 98 |
| rRNA (5S, 16S, 23S) | 2, 2, 2 | 2, 2, 2 | 2, 2, 2 | 2, 2, 2 |
| ncRNA | 18 | 17 | 18 | 38 |
| tRNA | 56 | 56 | 53 | 54 |
| Pseudogenes | 65 | 47 | 63 | 130 |
| ENA/GenBank accession number | GCA_903989455 | CGA_905187425 | GCA_903989465 | GCA_903989475 |
| Reference | [32] | This study | [32] | [34] |
| X. euroxanthea | Xaj | |||||
|---|---|---|---|---|---|---|
| Protein/Gene Name | Label | GenBank Accession Number | CPBF 367 | CPBF 424T | CPBF 426 | CPBF 427 |
| Chemotaxis-related proteins: | ||||||
| Methyl-accepting chemotaxis protein | XCV1938 | CAJ23615.1 | ● * | ● | ● | ○ |
| Chemotaxis protein | XAC3768 | AAM38611.1 | ● | ● | ● | ○ |
| Non-fimbrial adhesins: | ||||||
| Filamentous hemagglutinin-related protein (fhaB1) | FhaB1 | CAJ23537.1 | ● | ○ | ○ | ● |
| Filamentous hemagglutinin-related protein (fhaB2) | FhaB2 | CAJ23538.1 | ● | ○ | ○ | ● |
| Type II secretion system (T2SS): | ||||||
| General secretion pathway protein XpsN (xpsN) | XpsN | WP_011035909.1 | ● | ● | ● | ○ |
| Extracellular enzymes: | ||||||
| Xylosidase/arabinosidase (xylB) | XylB.1 | WP_011036375.1 | ○ | ● | ○ | ● |
| Xylosidase/arabinosidase (xsa) | Xsa.1 | WP_011037540.1 | ○ | ○ | ○ | ● |
| Xylosidase/arabinosidase (xylB) | XylB.2 | WP_011039174.1 | ○ | ● | ○ | ● |
| Endoglucanase (bcsZ) | bcsZ | AAM38359.1 | ○ | ○ | ○ | ● |
| Polygalacturonase (pglA) | pglA | WP_011037410.1 | ○ | ○ | ○ | ● |
| Pectate lyase E (pelA) | pelA | WP_011035380.1 | ● | ● | ● | ○ |
| Degenerated pectate lyase (pel) | pel | AAM37225.1 | ○ | ○ | ○ | ● |
| Pectinesterase | XCC0121 | WP_011035379.1 | ● | ● | ● | ○ |
| Type IV pilus (T4P): | ||||||
| PilY1 protein (pilY1) | pilY1 | WP_011051753.1 | ○ | ● | ○ | ○ |
| PilX protein (pilX) | pilX | WP_011051754.1 | ○ | ● | ○ | ○ |
| PilW protein (pilW) | XAC2667 | WP_040107776.1 | ○ | ● | ○ | ○ |
| Pre-pilin leader sequence (pilV) | pilV | WP_011051756.1 | ○ | ● | ○ | ○ |
| Pre-pilin like leader sequence (fimT) | fimT | WP_011051757.1 | ○ | ● | ○ | ○ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, C.; Martins, L.; Teixeira, M.; Blom, J.; Pothier, J.F.; Fonseca, N.A.; Tavares, F. Comparative Genomics of Xanthomonas euroxanthea and Xanthomonas arboricola pv. juglandis Strains Isolated from a Single Walnut Host Tree. Microorganisms 2021, 9, 624. https://doi.org/10.3390/microorganisms9030624
Fernandes C, Martins L, Teixeira M, Blom J, Pothier JF, Fonseca NA, Tavares F. Comparative Genomics of Xanthomonas euroxanthea and Xanthomonas arboricola pv. juglandis Strains Isolated from a Single Walnut Host Tree. Microorganisms. 2021; 9(3):624. https://doi.org/10.3390/microorganisms9030624
Chicago/Turabian StyleFernandes, Camila, Leonor Martins, Miguel Teixeira, Jochen Blom, Joël F. Pothier, Nuno A. Fonseca, and Fernando Tavares. 2021. "Comparative Genomics of Xanthomonas euroxanthea and Xanthomonas arboricola pv. juglandis Strains Isolated from a Single Walnut Host Tree" Microorganisms 9, no. 3: 624. https://doi.org/10.3390/microorganisms9030624