
Genetic Transformation of Quercus ilex Somatic Embryos with a Gnk2-like Protein That Reveals a Putative Anti-Oomycete Action

 ,
, 
 and
and
Abstract
:1. Introduction
2. Results
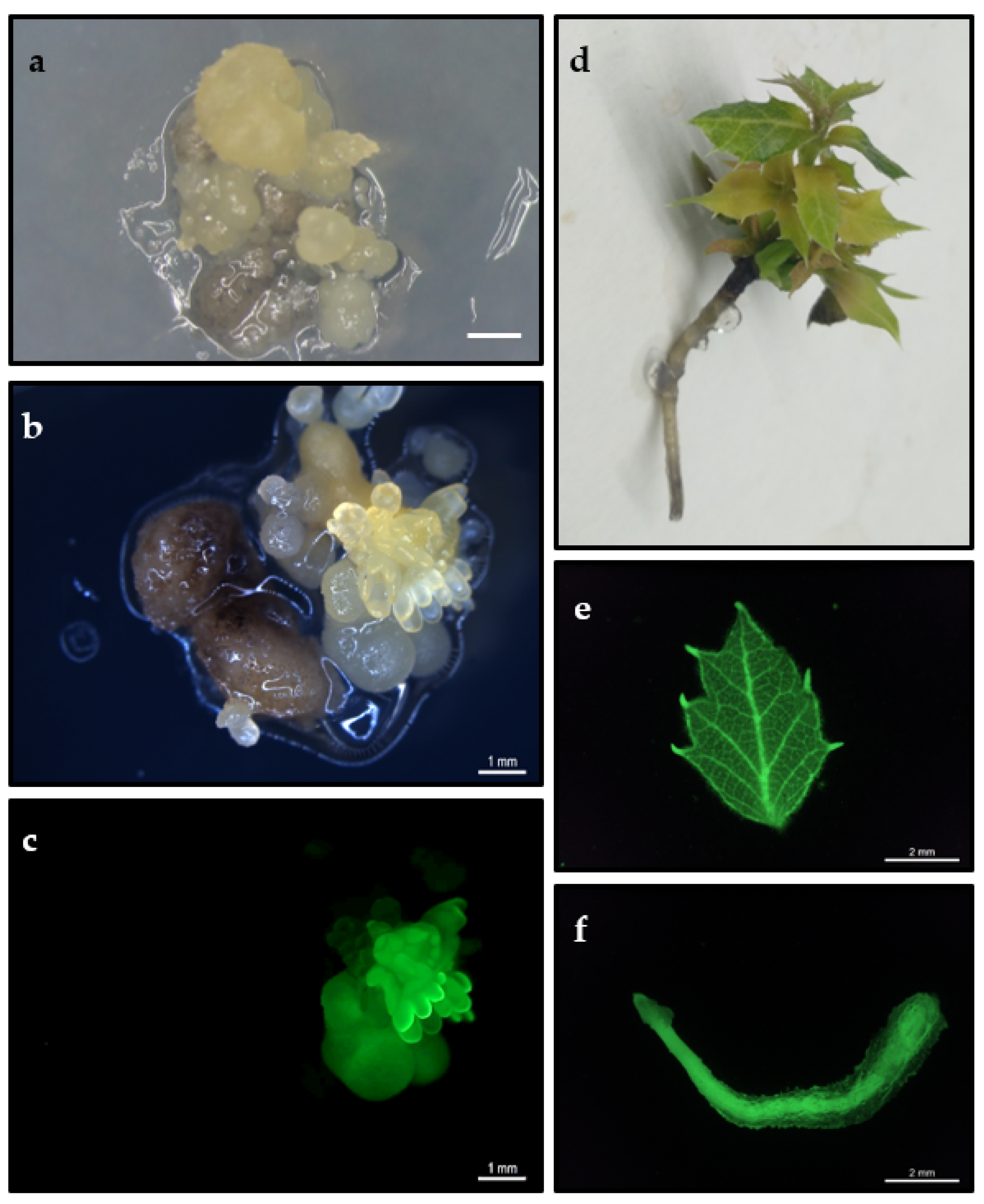
2.1. Genetic Transformation of Proembryogenic Masses
2.2. Maintenance of Transgenic Embryogenic Lines
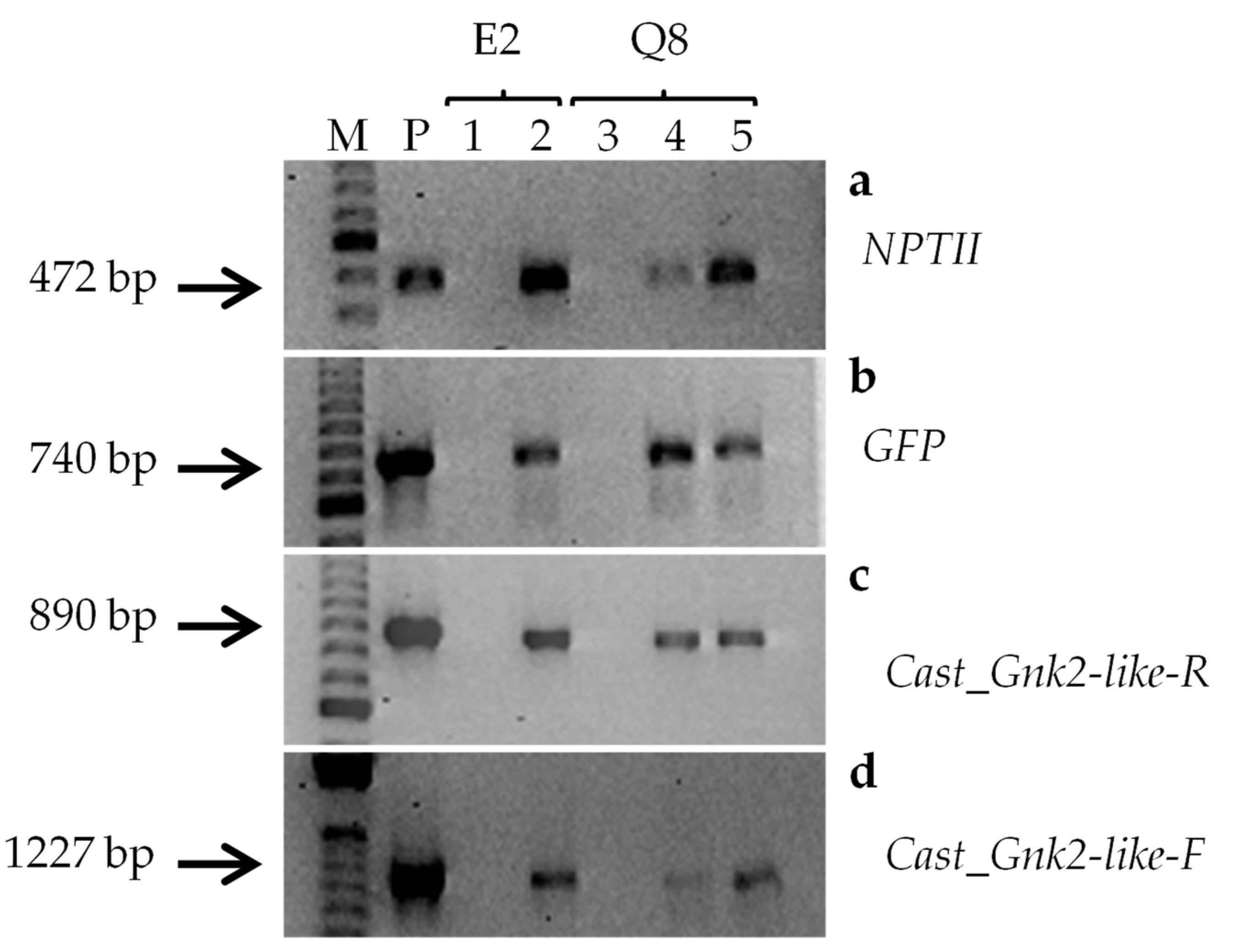
2.3. Molecular Analysis of Transgenic Embryogenic Lines
2.3.1. Gene Presence Analysis
2.3.2. Gene Number Copy Analysis
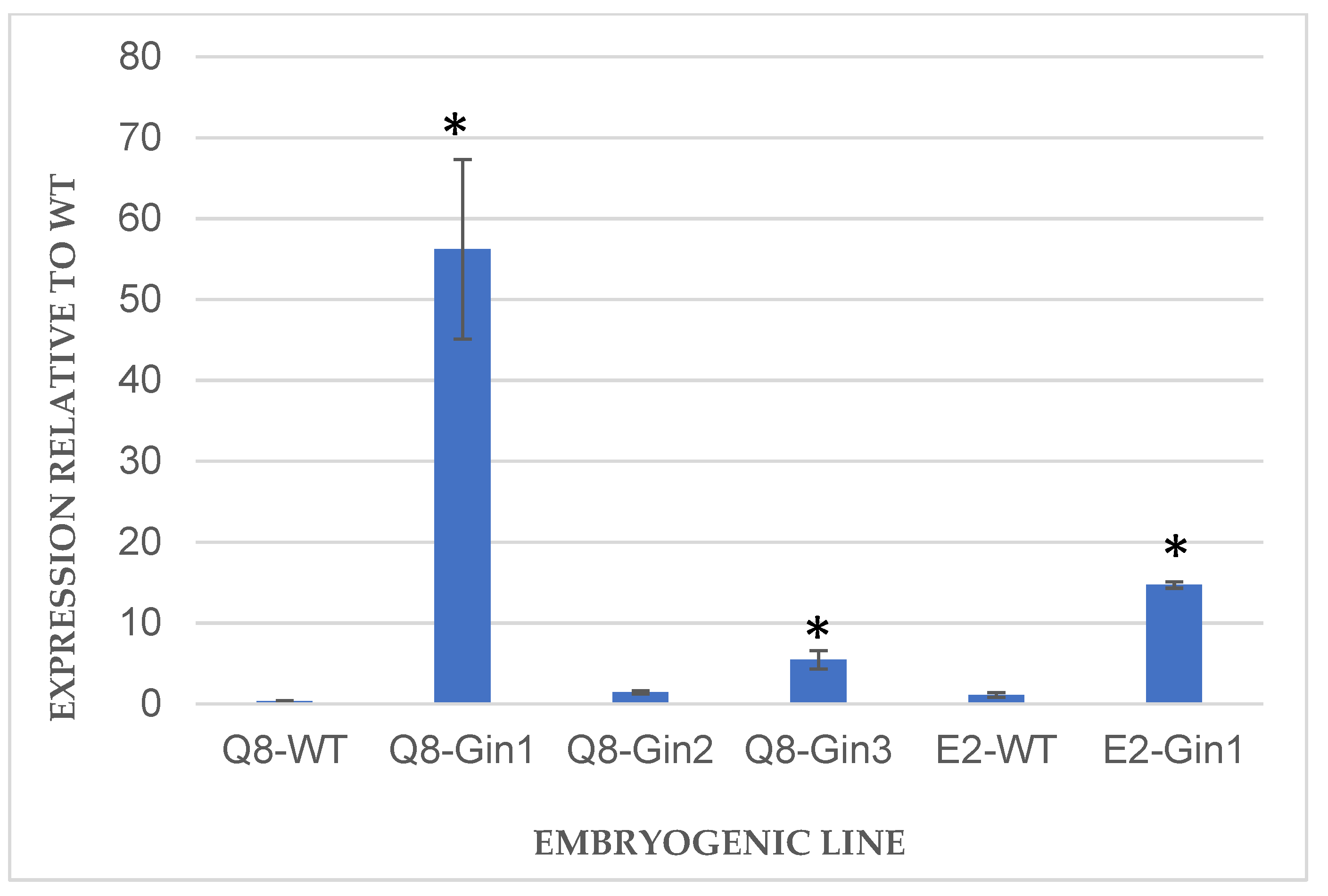
2.3.3. Gene Expression Analysis
2.4. Plant Regeneration from Transgenic Somatic Embryos
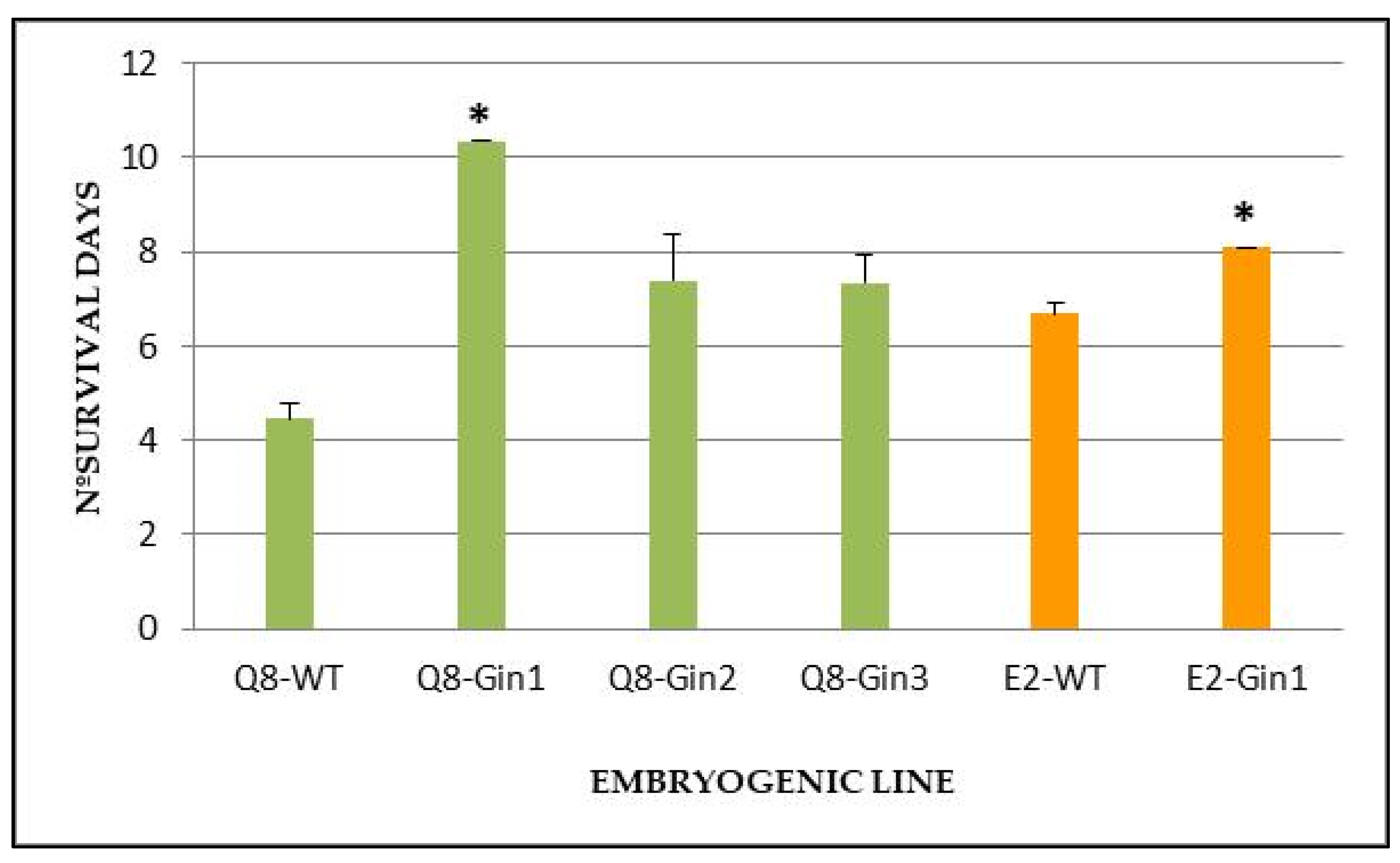
2.5. In Vitro Tolerance Assay
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Binary Plasmid Construction and Transformation into Agrobacterium Strain
4.3. Transformation Procedures
4.4. Selection and Proliferation of Putative Transgenic Lines
4.5. Molecular Confirmation of Transformation
4.5.1. Gene Presence Analysis
4.5.2. Gene Number Copy Analysis
4.5.3. Gene Expression Analysis
4.6. Regeneration of Transgenic Plants
4.7. Plant Pathogenicity Assay
4.7.1. Oomycete Strain and Sporangium and Zoospore Preparation
4.7.2. In Vitro Tolerance Assay
4.8. Data Handling and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stenlid, J.; Oliva, J.; Boberg, J.B.; Hopkins, A.J.M. Emerging Diseases in European Forest Ecosystems and Responses in Society. Forests 2011, 2, 486–504. [Google Scholar] [CrossRef]
- Sicard, P.; Augustaitis, A.; Belyazid, S.; Calfapietra, C.; de Marco, A.; Fenn, M.; Bytnerowicz, A.; Grulke, N.; He, S.; Matyssek, R.; et al. Global topics and novel approaches in the study of air pollution, climate change and forest ecosystems. Environ. Pollut. 2016, 213, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Haavik, L.J.; Billings, S.A.; Guldin, J.M.; Stephen, F.M. Emergent insects, pathogens and drought shape changing patterns in oak decline in North America and Europe. For. Ecol. Manag. 2015, 354, 190–205. [Google Scholar] [CrossRef]
- Natalini, F.; Alejano, R.; Vázquez-Piqué, J.; Cañellas, I.; Gea-Izquierdo, G. The role of climate change in the widespread mortality of holm oak in open woodlands of Southwestern Spain. Dendrochronologia 2016, 38, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Ruiz Gomez, F.J. Study of the Interaction between Root Rot Oomycetes and Quercus ilex L. Ph.D. Thesis, Universidad de Córdoba, Córdoba, Spain, 2018. [Google Scholar]
- Guzmán-Álvarez, J.R. The image of a tamed landscape: Dehesa through History in Spain. Cult. Hist. Digit. J. 2016, 5, e003. [Google Scholar] [CrossRef]
- Corcobado, T.; Cubera, E.; Moreno, G.; Solla, A. Quercus ilex forests are influenced by annual variations in water table, soil water deficit and fine root loss caused by Phytophthora cinnamomi. Agric. For. Meteorol. 2013, 169, 92–99. [Google Scholar] [CrossRef]
- Kamoun, S.; Furzer, O.; Jones, J.D.G.; Judelson, H.S.; Ali, G.S.; Dalio, R.J.; Roy, S.G.; Schena, L.; Zambounis, A.; Panabières, F.; et al. The Top 10 oomycete pathogens in molecular plant pathology: Top 10 oomycete plant pathogens. Mol. Plant Pathol. 2015, 16, 413–434. [Google Scholar] [CrossRef]
- Martínez, M.T.; Arrillaga, I.; Sales, E.; Pérez-Oliver, M.A.; González-Mas, M.C.; Corredoira, E. Micropropagation, characterization and conservation of Phytophthora cinnamomi-tolerant holm oak mature trees. Forest 2021, 12, 1634. [Google Scholar] [CrossRef]
- Martínez, M.T.; San-José, M.C.; Arrillaga, I.; Cano, V.; Morcillo, M.; Cernadas, M.J.; Corredoira, E. Holm oak somatic embryogenesis: Current status and future perspectives. Front. Plant Sci. 2019, 10, 239. [Google Scholar] [CrossRef] [Green Version]
- Merkle, S.A.; Dean, J.F.D. Forest tree biotechnology. Curr. Opin. Biotechnol. 2000, 11, 298–302. [Google Scholar] [CrossRef]
- Giri, C.C.; Shyamkumar, B.; Anjaneyulu, C. Progress in tissue culture, genetic transformation and applications of biotechnology to trees: An overview. Trees 2004, 18, 115–135. [Google Scholar] [CrossRef]
- Guan, Y.; Li, S.-G.; Fan, X.-F.; Su, Z.-H. Application of Somatic Embryogenesis in Woody Plants. Front. Plant Sci. 2016, 7, 938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parmar, N.; Singh, K.; Sharma, D.; Singh, L.; Kumar, P.; Nanjundan, J.; Khan, Y.J.; Chauhan, D.K.; Thakur, A.K. Genetic engineering strategies for biotic and abiotic stress tolerance and quality enhancement in horticultural crops: A comprehensive review. 3 Biotech. 2017, 7, 239. [Google Scholar] [CrossRef] [PubMed]
- Corredoira, E.; Merkle, S.A.; Martínez, M.T.; Toribio, M.; Canhoto, J.M.; Correia, S.I.; Ballester, A.; Vieitez, A.M. Non-zygotic embryogenesis in hardwood species. Crit. Rev. Plant Sci. 2019, 38, 29–97. [Google Scholar] [CrossRef]
- Corredoira, E.; Valladares, S.; Allona, I.; Aragoncillo, C.; Vieitez, A.M.; Ballester, A. Genetic Transformation of European Chestnut Somatic Embryos with a Native Thaumatin-Like Protein (CsTL1) Gene Isolated from Castanea sativa Seeds. Tree Physiol. 2012, 32, 1389–1402. [Google Scholar] [CrossRef] [Green Version]
- Mallón, R.; Valladares, S.; Corredoira, E.; Vieitez, A.M.; Vidal, N. Overexpression of the Chestnut CsTL1 Gene Coding for a Thaumatin-like Protein in Somatic Embryos of Quercus robur. Plant Cell Tissue Organ Cult. 2013, 116, 141–151. [Google Scholar] [CrossRef]
- Cano, V.; Martínez, M.T.; San José, M.C.; Couselo, J.L.; Varas, E.; Bouza-Morcillo, L.; Toribio, M.; Corredoira, E. Regeneration of Transgenic Plants by Agrobacterium-Mediated Transformation of Quercus ilex L. Somatic Embryos with the Gene CsTL1. New For. 2020, 51, 1003–1021. [Google Scholar] [CrossRef]
- Cano, V.; Martínez, M.T.; Couselo, J.L.; Varas, E.; Vieitez, F.J.; Corredoira, E. Efficient Transformation of Somatic Embryos and Regeneration of Cork Oak Plantlets with a Gene (CsTL1) Encoding a Chestnut Thaumatin-Like Protein. Int. J. Mol. Sci. 2021, 22, 1757. [Google Scholar] [CrossRef]
- Available online: http://www.hardwoodgenomics.org (accessed on 22 November 2021).
- Serrazina, S.; Santos, C.; Machado, H.; Pesquita, C.; Vicentini, R.; Pais, M.S.; Sebastiana, M.; Costa, R. Castanea root transcriptome in response to Phytophthora cinnamomi challenge. Tree Genet. Genomes 2015, 11, 6. [Google Scholar] [CrossRef]
- Santos, C.; Zhebentyayeva, T.; Serrazina, S.; Nelson, C.D.; Costa, R. Development and characterization of EST-SSR markers for mapping reaction to Phytophthora cinnamomi in Castanea spp. Sci. Hortic. 2015, 194, 181–187. [Google Scholar] [CrossRef]
- Santos, C.; Duarte, S.; Tedesco, S.; Fevereiro, P.; Costa, R.L. Expression Profiling of Castanea Genes during Resistant and Susceptible Interactions with the Oomycete Pathogen Phytophthora cinnamomi Reveal Possible Mechanisms of Immunity. Front. Plant Sci. 2017, 8, 515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyakawa, T.; Hatano, K.-I.; Miyauchi, Y.; Suwa, Y.-I.; Sawano, Y.; Tanokura, M. A Secreted Protein with Plant-Specific Cysteine-Rich Motif Functions as a Mannose-Binding Lectin That Exhibits Antifungal Activity. Plant Physiol. 2014, 166, 766–778. [Google Scholar] [CrossRef] [Green Version]
- Mélida, H.; Sandoval-Sierra, J.V.; Diéguez-Uribeondo, J.; Bulone, V. Analyses of Extracellular Carbohydrates in Oomycetes Unveil the Existence of Three Different Cell Wall Types. Eukaryot. Cell 2013, 12, 194–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, K.D.; Layne, D.R.; Scorza, R.; Schnabel, G. Gastrodia anti-fungal protein from the orchid Gastrodia elata confers disease resistance to root pathogens in transgenic tobacco. Planta 2006, 224, 1373. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Cai, C.; Skokut, M.; Kosegi, B.D.; Petolino, J.F. Quantitative real-time PCR as a screening tool for estimating transgene copy number in WHISKERS™-derived transgenic maize. Plant Cell Rep. 2002, 20, 948–954. [Google Scholar] [CrossRef]
- FAO; UNEP. The State of the World’s Forests 2020: Forests, Biodiversity and People; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Wang, H.; Ng, T.B. Ginkbilobin, a novel antifungal protein from Ginkgo biloba seeds with sequence similarity to embryo-abundant protein. Biochem. Biophys. Res. Commun. 2000, 279, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Yuan, S.S.; Jiang, L.L.; Ye, X.J.; Ng, T.B.; Wu, Z.J. Plant antifungal proteins and their applications in agriculture. Appl. Microbiol. Biotechnol. 2015, 99, 4961–4981. [Google Scholar] [CrossRef]
- Castro, M.S.; Fontes, W. Plant defense and antimicrobial peptides. Protein Pept. Lett. 2005, 12, 13–18. [Google Scholar] [CrossRef]
- Pelegrini, P.B.; Franco, O.L. Plant gamma-thionins: Novel insights on the mechanism of action of a multi-functional class of defense proteins. Int. J. Biochem. Cell 2005, 37, 2239–2253. [Google Scholar] [CrossRef]
- Stotz, H.U.; Thomson, J.; Wang, Y. Plant defensins. Plant Signal. Behav. 2009, 4, 1010–1012. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Tian, H.; Wang, Y.; Guo, A. Overexpression of a Novel Antifungal Protein Gene GNK2-1 Results in Elevated Resistance of Transgenic Cucumber to Fusarium oxysporum. Chin. Bull. Bot. 2010, 45, 411–418. [Google Scholar] [CrossRef]
- McGuigan, L.; Fernandes, P.; Oakes, A.; Stewart, K.; Powell, W. Transformation of American Chestnut (Castanea dentata (Marsh.) Borkh) Using RITA® Temporary Immersion Bioreactors and We Vitro Containers. Forests 2020, 11, 1196. [Google Scholar] [CrossRef]
- Ribas, A.F.; Dechamp, E.; Champion, A.; Bertrand, B.; Combes, M.C.; Verdeil, L.; Lapeyre, F.; Lashermes, P.; Etienne, P. Agrobacterium-mediated genetic transformation of Coffea arabica (L.) is greatly enhanced by using established embryogenic callus cultures. BMC Plant Biol. 2011, 11, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, S.; Mahon, E.L.; MacKay, H.A.; Rottmann, W.H.; Strauss, S.H.; Pijut, P.M.; Powell, W.A.; Coffey, V.; Lu, H.; Mansfield, S.D.; et al. Genetic engineering of trees: Progress and new horizons. In Vitro Cell. Dev. Biol. Plant 2018, 54, 341–376. [Google Scholar] [CrossRef]
- Dhekney, S.A.; Li, Z.T.; Dutt, M.; Gray, D.J. Agrobacterium-mediated transformation of embryogenic cultures and plant regeneration in Vitis rotundifolia Michx. (muscadine grape). Plant Cell Rep. 2008, 27, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Andrade, G.M.; Nairn, C.J.; Le, H.T.; Merkle, S.A. Sexually mature transgenic American chestnut trees via embryogenic suspension-based transformation. Plant Cell Rep. 2009, 28, 1385–1397. [Google Scholar] [CrossRef]
- Corredoira, E.; San-José, M.C.; Vieitez, A.M.; Ballester, A. Improving genetic transformation of European chestnut and cryopreservation of transgenic lines. Plant Cell Tissue Organ Cult. 2007, 91, 281–288. [Google Scholar] [CrossRef] [Green Version]
- Leroy, T.; Henry, A.M.; Royer, M.; Altosaar, I.; Frutos, R.; Duris, D.; Philippe, R. Genetically modified coffee plants expressing the Bacillus thuringiensis cry 1Ac gene for resistance to leaf miner. Plant Cell Rep. 2000, 19, 382–385. [Google Scholar] [CrossRef]
- Yancheva, S.D.; Shlizerman, L.A.; Golubowicz, S.; Yabloviz, Z.; Perl, A.; Hanania, U.; Flaishman, M.A. The use of green fluorescent protein (GFP) improves Agrobacterium-mediated transformation of “Spadona” pear (Pyrus communis L.). Plant Cell Rep. 2006, 25, 183–189. [Google Scholar] [CrossRef]
- Palomo-Ríos, E.; Cerezo, S.; Mercado, J.A.; Pliego-Alfaro, F. Agrobacterium-mediated transformation of avocado (Persea americana Mill.) somatic embryos with fluorescent marker genes and optimization of transgenic plant recovery. Plant Cell Tissue Organ Cult. 2017, 128, 447–455. [Google Scholar] [CrossRef]
- Leclercq, J.; Lardet, L.; Martin, F.; Chapuset, T.; Oliver, G.; Montoro, P. The green fluorescent protein as an efficient selection marker for Agrobacterium tumefaciens-mediated transformation in Hevea brasiliensis (Müll. Arg). Plant Cell Rep. 2010, 29, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Mallón, R.; Vieitez, A.M.; Vidal, N. High-efficiency Agrobacterium-mediated transformation in Quercus robur: Selection by use of a temporary immersion system and assessment by quantitative PCR. Plant Cell Tiss Organ Cult. 2013, 114, 171–185. [Google Scholar] [CrossRef] [Green Version]
- Matzke, A.J.; Matzke, M.A. Position effects and epigenetic silencing of plant transgenes. Curr. Opin. Plant Biol. 1998, 1, 142–148. [Google Scholar] [CrossRef]
- Raveendar, S.; Premkumar, A.; Sasikumar, S.; Ignacimuthu, S.; Agastian, P. Transgene and its expression in transgenic plants: A review. Plant Cell Biotechnol. Mol. Biol. 2007, 8, 105–112. [Google Scholar]
- Narváez, I.; Pliego Prieto, C.; Palomo-Ríos, E.; Fresta, L.; Jiménez-Díaz, R.M.; Trapero-Casas, J.L.; Lopez-Herrera, C.; Arjona-Lopez, J.M.; Mercado, J.A.; Pliego-Alfaro, F. Heterologous Expression of the AtNPR1 Gene in Olive and Its Effects on Fungal Tolerance. Front. Plant Sci. 2020, 11, 308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutt, M.; Grosser, J.W. An embryogenic suspension cell culture system for D-mediated transformation of citrus. Plant Cell Rep. 2010, 29, 1251–1260. [Google Scholar] [CrossRef]
- Martínez, M.T.; San-José, M.C.; Vieitez, A.M.; Cernadas, M.J.; Ballester, A.; Corredoira, E. Propagation of mature Quercus ilex L. (holm oak) trees by somatic embryogenesis. Plant Cell Tissue Organ Cult. 2017, 131, 321–333. [Google Scholar] [CrossRef]
- Flavell, R.B. Inactivation of gene expression in plants as a consequence of specific sequence duplication. Proc. Natl. Acad. Sci. USA 1994, 91, 3490–3496. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Rong, W.; Qi, L.; Li, J.; Wei, X.; Zhang, Z. Isolation and characterization of a novel wheat cysteine-rich receptor-like kinase gene induced by Rhizoctonia cerealis. Sci. Rep. 2013, 3, 3021. [Google Scholar] [CrossRef] [Green Version]
- Kimura, S.; Hunter, K.; Vaahtera, L.; Tran, H.C.; Citterico, M.; Vaattovaara, A.; Rokka, A.; Stolze, S.C.; Harzen, A.; Meißner, L.; et al. CRK2 and C-terminal Phosphorylation of NADPH Oxidase RBOHD Regulate Reactive Oxygen Species Production in Arabidopsis. Plant Cell 2020, 32, 1063–1080. [Google Scholar] [CrossRef] [Green Version]
- Mou, S.; Meng, Q.; Gao, F.; Zhang, T.; He, W.; Guan, D.; He, S. A cysteine-rich receptor-like protein kinase CaCKR5 modulates immune response against Ralstonia solanacearum infection in pepper. BMC Plant Biol. 2021, 21, 382. [Google Scholar] [CrossRef] [PubMed]
- Barra-Jiménez, A.; Blasco, M.; Ruiz-Galea, M.; Celestino, C.; Alegre, J.; Arrillaga, I.; Toribio, M. Cloning mature holm oak trees by somatic embryogenesis. Trees 2014, 28, 657–667. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrand, A.C. Medium and techniques for induction of growth of monocotyledonous and dicotyledonous plant cell culture. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hood, E.E.; Gelvin, S.B.; Melchers, L.S.; Hoekema, A. New Agrobacterium helper plasmids for gene transfer to plants. Transgenic Res. 1993, 2, 208–218. [Google Scholar] [CrossRef]
- Xu, R.; Li, Q.Q. Protocol: Streamline cloning of genes into binary vectors in Agrobacterium via the Gateway®TOPO vector system. Plant Methods 2008, 4, 4. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Nueva York, NY, USA, 1989. [Google Scholar]
- Chen, S.C.; Cannon, C.H.; Kua, C.S.; Liu, J.-J.; Galbraith, D.W. Genome size variation in the Fagaceae and its implications for trees. Tree Genet. Genomes 2014, 10, 977–988. [Google Scholar] [CrossRef]
- Kotrade, P.; Sehr, E.M.; Wischnitzki, E.; Brüggemann, W. Comparative transcriptomics-based selection of suitable reference genes for normalization of RT-qPCR experiments in drought-stressed leaves of three European Quercus species. Tree Genet. Genomes 2019, 15, 38. [Google Scholar] [CrossRef]
- Marum, L.; Miguel, A.; Ricardo, C.P.; Miguel, C. Reference Gene Selection for Quantitative Real-time PCR Normalization in Quercus suber. PLoS ONE 2021, 7, e35113. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Gresshoff, P.M.; Doy, C.H. Development and differentiation of haploid Lycopersicon esculentum. Planta 1972, 107, 161–170. [Google Scholar] [CrossRef] [PubMed]
- Corcobado, T.; Cubera, E.; Juárez, E.; Moreno, G.; Solla, A. Drought events determine performance of Quercus ilex seedlings and increase their susceptibility to Phytophthora cinnamomi. Agric. For. Meteorol. 2014, 192–193, 1–8. [Google Scholar] [CrossRef]
- Stevens, R.B. Mycology Guidebook; University of Washington Press: Seattle, WA, USA, 1974; 703p. [Google Scholar]
- Rial, C.; Barros, A.; Salinero, C.; Mansilla, J.P.; Pintos, C.; Cuenca, B. Variaciones en la susceptibilidad a Phytophthora cinnamomi de diferentes clones de castaño: Comparación de dos métodos de inoculación. Cuad. Soc. Esp. Cienc. For. 2008, 26, 171–176. [Google Scholar] [CrossRef]
- Chen, D.W.; Zentmyer, G.A. Production of sporangia by Phytophthora cinnamomi in axenic culture. Mycologia 1970, 62, 397–402. [Google Scholar] [CrossRef]
- Cuenca, B.; Ocaña, L.; Salinero, M.C.; Pintos, C.; Mansilla, J.P.; Rial, C. Selection of Castanea sativa Mill., for resistance to Phytophthora cinnamomi: Micropropagation and testing of selected clones. Acta Hortic. 2010, 866, 111–119. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Embryogenic Line | Kanamycin Resistant Explants (%) 1 | Transformation Efficiency (%) 2 | Selection Efficiency (%) 3 |
|---|---|---|---|
| Q8 | 5.0 ± 2.7 | 2.5 ± 1.7 | 50.0 |
| E2 | 3.3 ± 1.4 | 2.5 ± 1.3 | 75.0 |
| Q10-16 | 0.0 ± 0.0 | 0.0 ± 0.0 | - |
| E00 | 0.0 ± 0.0 | 0.0 ± 0.0 | - |
| χ2 | p < 0.05 | ns | - |
| Line | CT Mean | CT Standard Deviation | Estimated Copy Number 1 |
|---|---|---|---|
| Q8-WT | 27.27 | 0.97 | 0 |
| Q8-Gin1 | 21.55 | 0.61 | 1 |
| Q8-Gin2 | 20.51 | 0.27 | 2 |
| E2-WT | 27.35 | 1.35 | 0 |
| E2-Gin1 | 21.69 | 0.41 | 1 |
| Line | Only Root | Conversion (Shoot + Root) | ||||
|---|---|---|---|---|---|---|
| (%) | RL (mm) | (%) | RL (mm) | SL (mm) | NL | |
| Lines Q8 | ||||||
| Q8-WT | 58.0 ± 6.8 | 52.5 ± 5.9 | 38.9 ± 5.1 | 54.9 ± 7.5 | 10.8 ± 0.9 | 7.6 ± 0.8 |
| Q8-Gin1 | 33.3 ± 14.7 | 88.8 ± 9.9 | 52.8 ± 16.4 | 69.7 ± 1.8 | 11.6 ± 1.2 | 7.9 ± 0.2 |
| Q8-Gin2 | 11.1 ± 3.2 | 105.0 ± 18.4 | 69.5 ± 10.0 | 78.3 ± 7.4 | 12.1 ± 0.9 | 9.5 ± 0.7 |
| Q8-Gin3 | 47.2 ± 12.1 | 58.9 ± 7.4 | 30.6 ± 4.7 | 42.2 ± 8.5 | 10.1 ± 1.9 | 9.5 ± 2.1 |
| ANOVA I | ns | ns | ns | p < 0.05 | ns | ns |
| Lines E2 | ||||||
| E2-WT | 55.6 ± 10.1 | 48.5 ± 5.6 | 38.9 ± 10.1 | 71.1 ± 13.1 | 16.7 ± 1.8 | 8.0 ± 1.3 |
| E2-Gin1 | 8.3 ± 3.4 | 100.0 ± 30.1 | 22.2 ± 8.5 | 62.3 ± 9.2 | 15.5 ± 0.4 | 4.8 ± 0.8 |
| ANOVA I | p < 0.05 | ns | ns | ns | ns | ns |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Serrazina, S.; Martínez, M.T.; Cano, V.; Malhó, R.; Costa, R.L.; Corredoira, E. Genetic Transformation of Quercus ilex Somatic Embryos with a Gnk2-like Protein That Reveals a Putative Anti-Oomycete Action. Plants 2022, 11, 304. https://doi.org/10.3390/plants11030304
Serrazina S, Martínez MT, Cano V, Malhó R, Costa RL, Corredoira E. Genetic Transformation of Quercus ilex Somatic Embryos with a Gnk2-like Protein That Reveals a Putative Anti-Oomycete Action. Plants. 2022; 11(3):304. https://doi.org/10.3390/plants11030304
Chicago/Turabian StyleSerrazina, Susana, Mª Teresa Martínez, Vanesa Cano, Rui Malhó, Rita Lourenço Costa, and Elena Corredoira. 2022. "Genetic Transformation of Quercus ilex Somatic Embryos with a Gnk2-like Protein That Reveals a Putative Anti-Oomycete Action" Plants 11, no. 3: 304. https://doi.org/10.3390/plants11030304