
Evaluation of Commercial Myxomatosis Vaccines against Recombinant Myxoma Virus (ha-MYXV) in Iberian Hare and Wild Rabbit

 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
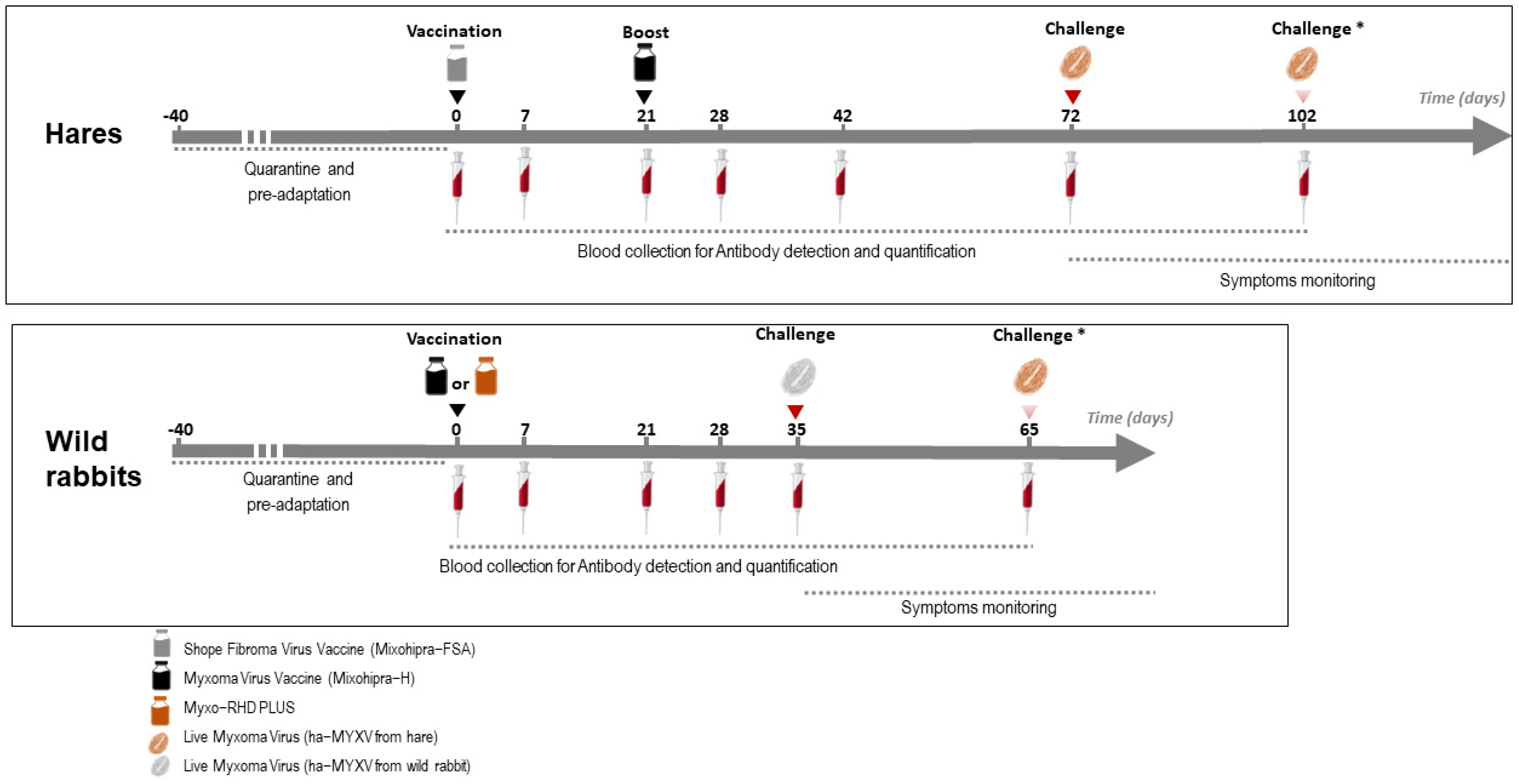
2.1. Origin of Animals and Pre-Adaptation to Captivity Conditions
2.2. Vaccines
2.3. Experimental Design
- (i)
- H-G1: a negative control group of three animals (#231, #232 and #233), MYXV seronegative and non-vaccinated;
- (ii)
- H-G2: a group of three animals (#076, #077 and #078), MYXV seronegative, vaccinated with a 2.90 × 104 ffu (focus-forming units) dose of Mixohipra-FSA vaccine at the start of the study and 21 days later with a 1.95 × 104 ffu dose of Mixohipra-H vaccine;
- (iii)
- H-G3: a group of three animals (#042, #043 and #044), MYXV seronegative, vaccinated with a 2.90 × 105 ffu dose of Mixohipra-FSA vaccine at the start of the study and 21 days later a 1.95 × 105 dose of Mixohipra-H vaccine;
- (iv)
- H-G4: a positive control animal (#10), MYXV seropositive, collected from the field after natural recovery from myxomatosis.
- (i)
- R-G1: a negative control group of four animals (#449, #451, #000 and #001), MYXV seronegative and non-vaccinated;
- (ii)
- R-G2: a group of three animals (#442, #444 and #445), MYXV seronegative, vaccinated with a 1.95 × 104 ffu dose of Mixohipra-H vaccine at the start of the study;
- (iii)
- R-G3: a group of three animals (#446, #447 and #448), MYXV seronegative, vaccinated with a single dose of Nobivac Myxo-RHD PLUS vaccine (103.0–105.8 ffu of each two vectors) at the start of the study.
2.4. Sampling and Blood Analyses
2.5. Serological Analyses
2.6. Viruses
2.7. Virus Detection by qPCR
2.8. Challenge
2.9. Clinical Signs Monitoring
2.10. Necropsy and Histopathology
2.11. Ethical and Legal Framework
3. Results
3.1. Humoral Immune Response to Vaccination
3.2. Hematologic and Biochemical Analyses of Blood Samples Obtained during Vaccination
3.3. Clinical Course after Challenge
3.4. Virus Presence in Drinking Water and Faeces
3.5. Necropsy, Histopathology and Virus Loads in Tissues
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Águeda-Pinto, A.; De Matos, A.L.; Abrantes, M.; Kraberger, S.; Risalde, M.A.; Gortázar, C.; McFadden, G.; Varsani, A.; Esteves, P.J. Genetic Characterization of a Recombinant Myxoma Virus in the Iberian Hare (Lepus granatensis). Viruses 2019, 11, 530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalton, K.P.; Martín, J.M.; Nicieza, I.; Podadera, A.; De Llano, D.; Casais, R.; Gimenez, S.; Badiola, I.; Agüero, M.; Duran, M.; et al. Myxoma virusjumps species to the Iberian hare. Transbound. Emerg. Dis. 2019, 66, 2218–2226. [Google Scholar] [CrossRef]
- Carvalho, C.L.; Dos Santos, F.A.A.; Monteiro, M.; Carvalho, P.; Mendonça, P.; Duarte, M.D. First cases of myxomatosis in Iberian hares (Lepus granatensis) in Portugal. Vet. Rec. Case Rep. 2020, 8, e001044. [Google Scholar] [CrossRef]
- Dos Santos, F.A.A.; Monteiro, M.; Pinto, A.; Carvalho, C.L.; Peleteiro, M.C.; Carvalho, P.; Mendonça, P.; Carvalho, T.; Duarte, M.D. First description of a herpesvirus infection in genus Lepus. PLoS ONE 2020, 15, e0231795. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.D.; Carvalho, C.L.; Dos Santos, F.A.; Monteiro, J.; Monteiro, M.; Carvalho, P.M.; Mendonça, P.; Santos, P.T.; Melo, P.C. The Health and Future of the Six Hare Species in Europe: A Closer Look at the Iberian Hare. In Lagomorpha Characteristics; IntechOpen: London, UK, 2020. [Google Scholar]
- García-Bocanegra, I.; Camacho-Sillero, L.; Caballero-Gómez, J.; Agüero, M.; Gómez-Guillamón, F.; Ruiz-Casas, J.M.; Díaz-Cao, J.M.; García, E.; Ruano, M.J.; de la Haza, R. Monitoring of emerging myxoma virus epidemics in Iberian hares (Lepus granatensis) in Spain, 2018–2020. Transbound. Emerg. Dis. 2020, 68, 1275–1282. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, F.A.; Carvalho, C.; Pinto, A.; Rai, R.; Monteiro, M.; Carvalho, P.; Mendonça, P.; Peleteiro, M.; Parra, F.; Duarte, M. Detection of recombinant Hare Myxoma Virus in wild rabbits (Oryctolagus cuniculus algirus). Viruses 2020, 12, 1127. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, F.A.A.; Carvalho, C.L.; Monteiro, M.; Carvalho, P.; Mendonça, P.; Peleteiro, M.D.C.; Duarte, M.D. Recombinant myxoma virus infection associated with high mortality in rabbit farming (Oryctolagus cuniculus). Transbound. Emerg. Dis. 2020, 68, 2616–2621. [Google Scholar] [CrossRef]
- Villafuerte, R.; Delibes-Mateos, M. Oryctolagus cuniculus. IUCN Red List 2019, 8235. [Google Scholar] [CrossRef]
- Soriguer, R.; Carro, F.; Lepus granatensis. IUCN Red List 2019: E.T41306A2953195. Available online: https://www.iucnredlist.org/species/41306/2953195 (accessed on 10 January 2022).
- Delibes-Mateos, M.; Ferreras, P.; Villafuerte, R. European rabbit population trends and associated factors: A review of the situation in the Iberian Peninsula. Mammal Rev. 2009, 39, 124–140. [Google Scholar] [CrossRef]
- Reemers, S.; Peeters, L.; Van Schijndel, J.; Bruton, B.; Sutton, D.; Van Der Waart, L.; Van De Zande, S. Novel Trivalent Vectored Vaccine for Control of Myxomatosis and Disease Caused by Classical and a New Genotype of Rabbit Haemorrhagic Disease Virus. Vaccines 2020, 8, 441. [Google Scholar] [CrossRef]
- Dos Santos, F.A.A.; Carvalho, C.L.; Peleteiro, M.C.; Gabriel, S.I.; Patrício, R.; Carvalho, J.; Cunha, M.V.; Duarte, M. Blood collection from the external jugular vein of Oryctolagus cuniculus algirus sedated with midazolam: Live sampling of a subspecies at risk. Wildl. Biol. 2019, 2019, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Smallwood, S.E.; Rahman, M.M.; Smith, D.W.; McFadden, G. Myxoma Virus: Propagation, Purification, Quantification, and Storage. Curr. Protoc. Microbiol. 2010, 17, 14A.1.1–14A.1.20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baer, A.; Kehn-Hall, K. Viral Concentration Determination Through Plaque Assays: Using Traditional and Novel Overlay Systems. J. Vis. Exp. 2014, e52065. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, F.A.A.; Carvalho, C.L.; Parra, F.; Dalton, K.P.; Peleteiro, M.C.; Duarte, M.D. A Quadruplex qPCR for Detection and Differentiation of Classic and Natural Recombinant Myxoma Virus Strains of Leporids. Int. J. Mol. Sci. 2021, 22, 12052. [Google Scholar] [CrossRef]
- dos Santos, F.A.; Carvalho, C.; Peleteiro, M.; Parra, F.; Duarte, M. A Versatile qPCR for Diagnosis of Leporid Gammaherpesvirus 5 Using Evagreen® or Taqman® Technologies. Viruses 2021, 13, 715. [Google Scholar] [CrossRef] [PubMed]
- Fenner, F.; Ratcliffe, F.N. Myxomatosis; Cambridge Universuty Press: London, UK, 1965. [Google Scholar]
- García-Bocanegra, I.; Camacho-Sillero, L.; Risalde, M.A.; Dalton, K.P.; Caballero-Gómez, J.; Agüero, M.; Zorrilla, I.; Gómez-Guillamón, F. First outbreak of myxomatosis in Iberian hares (Lepus granatensis). Transbound. Emerg. Dis. 2019, 66, 2204–2208. [Google Scholar] [CrossRef] [PubMed]
- Duarte, M.; Carvalho, C.L.; Santos, F.A.; Monteiro, J.; Gomes, J.; Alves, P.C.; Esteves, P.J.; Abrantes, J.; Lopes, A.M.; Monterroso, P.; et al. +Coelho2: Desenvolvimento e implementação de medidas práticas impulsionadoras da recuperação dos leporídeos silvestres em Portugal. In Relatório de Atividades na Época Venatória 2018–2019 e 2019–2020; Instituto Nacional de Investigação Agrária e Veterinária, I. P. (INIAV, I. P.): Oeiras, Portugal, 2020; Available online: https://www.iniav.pt/images/Noticias/Coelho2_Final-Report-web.pdf (accessed on 10 January 2022).
- Guitton, J.-S.; Devillard, S.; Guénézan, M.; Fouchet, D.; Pontier, D.; Marchandeau, S. Vaccination of free-living juvenile wild rabbits (Oryctolagus cuniculus) against myxomatosis improved their survival. Prev. Veter- Med. 2008, 84, 1–10. [Google Scholar] [CrossRef]
- Cabezas, S.; Calvete, C.; Moreno, S. Vaccination Success and Body Condition in the European Wild Rabbit: Applications for Conservation Strategies. J. Wildl. Manag. 2006, 70, 1125–1131. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, C.; Ramírez, E.; Castro, F.; Ferreras, P.; Alves, P.C.; Redpath, S.; Villafuerte, R. Field experimental vaccination campaigns against myxomatosis and their effectiveness in the wild. Vaccine 2009, 27, 6998–7002. [Google Scholar] [CrossRef]
- Marlier, D. Vaccination strategies against myxomavirus infections: Are we really doing the best? Tijdschr. Diergeneeskd. 2010, 135, 194–198. [Google Scholar]
- Kerr, P.J.; Merchant, J.C.; Silvers, L.; Hood, G.M.; Robinson, A.J. Monitoring the spread of myxoma virus in rabbit Oryctolagus cuniculus populations on the southern tablelands of New South Wales, Australia. II. Selection of a strain of virus for release. Epidemiology Infect. 2003, 130, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashton, L.V.; Quackenbush, S.L.; Castle, J.; Wilson, G.; McCoy, J.; Jordan, M.; MacNeill, A.L. Recombinant Myxoma Virus Expressing Walleye Dermal Sarcoma Virus or fC Is Attenuated in Rabbits. Viruses 2020, 12, 517. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.; Nicieza, I.; de Llano, D.; Gullón, J.; Inza, M.; Petralanda, M.; Arroita, Z.; Parra, F. Vaccine breaks: Outbreaks of myxomatosis on Spanish commercial rabbit farms. Veter- Microbiol. 2015, 178, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Manev, I.; Genova, K.; Lavazza, A.; Capucci, L. Humoral immune response to different routes of myxomatosis vaccine application. World Rabbit Sci. 2018, 26, 149–154. [Google Scholar] [CrossRef]
- Levin, C.; Perrin, H.; Combadiere, B. Tailored immunity by skin antigen-presenting cells. Hum. Vaccines Immunother. 2014, 11, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.C.; Jarrahian, C.; Zehrung, D.; Mitragotri, S.; Prausnitz, M.R. Delivery Systems for Intradermal Vaccination. Intradermal Immun. 2011, 351, 77–112. [Google Scholar] [CrossRef]
- Psikal, I.; Smid, B.; Rodak, L.; Valicek, L.; Bendova, J. Atypical myxomatosis-virus isolation, experimental infection of rabbits and restriction endonuclease analysis of the isolate. J. Veter- Med. Ser. B 2003, 50, 259–264. [Google Scholar] [CrossRef]
- Marshall, I.D.; Fenner, F. Studies in the epidemiology of infectious myxomatosis of rabbits: VII. The virulence of strains of myxoma virus recovered from Australian wild rabbits between 1951 and 1959. J. Hyg. 1960, 58, 485–488. [Google Scholar] [CrossRef] [Green Version]
- Kerr, P.; Perkins, H.; Inglis, B.; Stagg, R.; McLaughlin, E.; Collins, S.; Van Leeuwen, B. Expression of rabbit IL-4 by recombinant myxoma viruses enhances virulence and overcomes genetic resistance to myxomatosis. Virology 2004, 324, 117–128. [Google Scholar] [CrossRef] [Green Version]
- Kappler-Gratias, S.; Bucher, L.; Top, S.; Quentin-Froignant, C.; Desbois, N.; Bertagnoli, S.; Louison, M.; Monge, E.; Bousquet-Melou, A.; Lacroix, M.; et al. Antipoxvirus Activity Evaluation of Optimized Corroles Based on Development of Autofluorescent ANCHOR Myxoma Virus. ACS Infect. Dis. 2021, 7, 2370–2382. [Google Scholar] [CrossRef]
- Boutard, B.; Vankerckhove, S.; Markine-Goriaynoff, N.; Sarlet, M.; Desmecht, D.; McFadden, G.; Vanderplasschen, A.; Gillet, L. The α2,3-Sialyltransferase Encoded by Myxoma Virus Is a Virulence Factor that Contributes to Immunosuppression. PLoS ONE 2015, 10, e0118806. [Google Scholar] [CrossRef] [PubMed]
- Bárcena, J.; Morales, N.; Boga, A.; Parra, F.; Sa, M.; Lucientes, J.; Page, A.; Blasco, R.; Torres, J.M. Horizontal Transmissible Protection against Myxomatosis and Rabbit Hemorrhagic Disease by Using a Recombinant Myxoma Virus. J. Virol. 2000, 74, 1114–1123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- OIE. Terrestrial Manual Myxomatosis. In Lagomorpha; OIE: Paris, France, 2018; pp. 1371–1388. [Google Scholar]
- Kerr, P.J.; Cattadori, I.M.; Rogers, M.B.; Fitch, A.; Geber, A.; Liu, J.; Sim, D.G.; Boag, B.; Eden, J.-S.; Ghedin, E.; et al. Genomic and phenotypic characterization of myxoma virus from Great Britain reveals multiple evolutionary pathways distinct from those in Australia. PLoS Pathog. 2017, 13, e1006252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spibey, N.; McCabe, V.J.; Greenwood, N.M.; Jack, S.C.; Sutton, D.; van der Waart, L. Novel bivalent vectored vaccine for control of myxomatosis and rabbit haemorrhagic disease. Veter- Rec. 2012, 170, 309. [Google Scholar] [CrossRef] [Green Version]
- Guerin, J.-L.; Gelfi, J.; Camus, C.; Delverdier, M.; Whisstock, J.; Amardeihl, M.-F.; Py, R.; Bertagnoli, S.; Messud-Petit, F. Characterization and functional analysis of Serp3: A novel myxoma virus-encoded serpin involved in virulence. J. Gen. Virol. 2001, 82, 1407–1417. [Google Scholar] [CrossRef]
- Fenner, B.Y.F.; Marshall, I.D. A comparison of the virulence for european rabbits (Oryctolagus cuniculus) of strains of Myxoma virus recovered in the field in Australi, Europe and America. J. Hyg. 1957, 55, 149–191. [Google Scholar] [CrossRef] [Green Version]
- Fenner, F.; Woodroofe, G.M. Changes in the virulence and antigenic structure of strains of myxoma virus recovered from Australian wild rabbits between 1950 and 1964. Aust. J. Exp. Biol. Med. Sci. 1965, 43, 359–370. [Google Scholar] [CrossRef]
- Jeklova, E.; Leva, L.; Matiasovic, J.; Kovarcik, K.; Kudlackova, H.; Nevorankova, Z.; Psikal, I.; Faldyna, M. Characterisation of immunosuppression in rabbits after infection with myxoma virus. Veter- Microbiol. 2008, 129, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Sousa, M.; Silva, V.; Silva, A.; Silva, N.; Ribeiro, J.; Tejedor-Junco, M.T.; Capita, R.; Chenouf, N.S.; Alonso-Calleja, C.; Rodrigues, T.M.; et al. Staphylococci among Wild European Rabbits from the Azores: A Potential Zoonotic Issue? J. Food Prot. 2020, 83, 1110–1114. [Google Scholar] [CrossRef]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [Green Version]
- Barber, G.R.; Lauretta, J.; Saez, R. A Febrile Neutropenic Patient with Enterococcus gallinarum Sepsis Treated with Daptomycin and Gentamicin. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2007, 27, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fan, S.; Lin, X.; Liu, L.; Zheng, J.; Zhou, X.; Heep, A. The first case report of Enterococcus gallinarum meningitis in neonate. Medicine 2018, 97, e9875. [Google Scholar] [CrossRef] [PubMed]
- Roig, F.J.; González-Candelas, F.; Sanjuan, E.; Fouz, B.; Feil, E.J.; Llorens, C.; Baker-Austin, C.; Oliver, J.D.; Danin-Poleg, Y.; Gibas, C.J.; et al. Phylogeny of Vibrio vulnificus from the Analysis of the Core-Genome: Implications for Intra-Species Taxonomy. Front. Microbiol. 2018, 8, 2613. [Google Scholar] [CrossRef] [PubMed]
- Horseman, M.A.; Surani, S. A comprehensive review of Vibrio vulnificus: An important cause of severe sepsis and skin and soft-tissue infection. Int. J. Infect. Dis. 2011, 15, e157–e166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsokana, C.N.; Sokos, C.; Giannakopoulos, A.; Birtsas, P.; Valiakos, G.; Spyrou, V.; Athanasiou, L.V.; Burriel, A.R.; Billinis, C. European Brown hare (Lepus europaeus) as a source of emerging and re-emerging pathogens of Public Health importance: A review. Veter- Med. Sci. 2020, 6, 550–564. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Humoral Response | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Group | Hare ID | Vaccine | Technique | Day 0 | Day 7 | Day 21 | Vaccine | Day 28 | Day 42 | Day 72 |
| H-G1 | #231 | Not vac | iELISA | 0.5 | 0.23 | 0.16 | Not vac | 0.13 | 0.19 | 0.23 |
| IFT | <2 | <2 | <2 | <2 | <2 | <2 | ||||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | ||||
| #232 | Not vac | iELISA | 0.35 | 0.16 | 0.39 | Not vac | 0.29 | 0.15 | 0.28 | |
| IFT | <2 | <2 | <2 | <2 | <2 | <2 | ||||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | ||||
| #233 | Not vac | iELISA | 0.63 | 0.47 | 0.42 | Not vac | 0.22 | 0.22 | 0.08 | |
| IFT | <2 | <2 | <2 | <2 | <2 | <2 | ||||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | ||||
| H-G2 | #076 | 1× M-FSA | iELISA | 0.9 | 1.01 | 0.84 | 1× M-H | 0.59 | 0.59 | 0.18 |
| IFT | <2 | 3 | 3.5 | 4 | 4 | 4 | ||||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | ||||
| #077 | 1× M-FSA | iELISA | 0.31 | 0.81 | 1.55 | 1× M-H | 2.45 | 2.81 | - | |
| IFT | <2 | <2 | 4 | 4 | 5.5 | |||||
| SNT | <1/4 | <1/4 | <1/4 | 1/8 | 1/8 | |||||
| #078 | 1× M-FSA | iELISA | 0.96 | 0.49 | 0.4 | 1× M-H | 0.36 | 0.36 | 0.12 | |
| IFT | <2 | <2 | 3 | 3 | 3 | 3 | ||||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | ||||
| H-G3 | #042 | 10× M-FSA | iELISA | 0.6 | 0.36 | 0.25 | 10× M-H | 0.24 | 0.18 | 0.03 |
| IFT | <2 | <2 | 3 | 3 | 3 | 3 | ||||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | ||||
| #043 | 10× M-FSA | iELISA | 0.79 | 0.62 | 5.55 | 10× M-H | 5.05 | 5.5 | 5.8 | |
| IFT | <2 | 4.5 | 6.5 | 7 | 7 | 7.5 | ||||
| SNT | <1/4 | <1/4 | 1/16 | 1/16–1/32 | 1/16–1/32 | 1/16–1/32 | ||||
| #044 | 10× M-FSA | iELISA | 0.75 | 0.36 | 5.57 | 10× M-H | 6.87 | 6.7 | 7.1 | |
| IFT | <2 | 3.5 | 7 | 7.5 | 7 | 8.5 | ||||
| SNT | <1/4 | <1/4 | 1/16 | 1/16–1/32 | 1/16–1/32 | 1/16–1/32 | ||||
| H-G4 | #10 | Natural immunity | iELISA | 7.09 | Natural immunity | 7.1 | 7.19 | 7.28 | ||
| IFT | 9 | 9 | 9 | 9 | ||||||
| SNT | 1/128 | 1/128 | 1/128 | 1/128 | ||||||
| Humoral Response | ||||||||
|---|---|---|---|---|---|---|---|---|
| Group | Rabbits ID | Vaccine | Technique | Day 0 | Day 7 | Day 21 | Day 28 | Day 35 |
| R-G1 | #449 | Not vac | iELISA | 0.05 | 0.06 | 0.07 | 0.06 | 0.02 |
| IFT | <2 | <2 | <2 | <2 | <2 | |||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | |||
| #451 | Not vac | iELISA | 0.22 | 0.01 | 0.1 | 0.27 | 0.2 | |
| IFT | <2 | <2 | <2 | <2 | <2 | |||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | |||
| #000 | Not vac | iELISA | 0.13 | 0.09 | 0.15 | 0.18 | 0.31 | |
| IFT | <2 | <2 | <2 | <2 | <2 | |||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | |||
| #001 | Not vac | iELISA | 0.32 | 0.27 | 0.38 | 0.21 | 0.14 | |
| IFT | <2 | <2 | <2 | <2 | <2 | |||
| SNT | <1/4 | <1/4 | <1/4 | <1/4 | <1/4 | |||
| R-G2 | #442 | 1× M-H | iELISA | 0.01 | 0.05 | 1.79 | 2.33 | 4.11 |
| IFT | <2 | 4 | 6 | 7.5 | 9 | |||
| SNT | <1/4 | <1/4 | <1/4 | 1/8 | 1/32 | |||
| #444 | 1× M-H | iELISA | 0.04 | 0.18 | 1.1 | 1.04 | 1.72 | |
| IFT | <2 | 2 | 5.5 | 6 | 7 | |||
| SNT | <1/4 | <1/4 | <1/4 | 1/4 | 1/8 | |||
| #445 | 1× M-H | iELISA | 0.1 | 0.29 | 0.83 | 1.04 | 2.15 | |
| IFT | <2 | 3 | <2 | 6 | 8.5 | |||
| SNT | <1/4 | <1/4 | <1/4 | 1/4 | 1/8 | |||
| R-G3 | #446 | 1× M-RHD | iELISA | 0.31 | 0.36 | 2.3 | 3.05 | 4.78 |
| IFT | <2 | 3.5 | 6.5 | 8 | 9 | |||
| SNT | <1/4 | <1/4 | 1/8 | 1/16 | 1/64 | |||
| #447 | 1× M-RHD | iELISA | 0.35 | 0.21 | 5.32 | 5.42 | 5.39 | |
| IFT | <2 | 4.5 | 9.5 | 10 | 10 | |||
| SNT | <1/4 | 1/4 | 1/128 | 1/128 | 1/128 | |||
| #448 | 1× M-RHD | iELISA | 0.26 | 0.51 | 1.21 | 1.09 | 2.15 | |
| IFT | <2 | 4 | 5.5 | 6 | 7 | |||
| SNT | <1/4 | <1/4 | 1/4 | 1/8 | 1/8–1/16 | |||
| Days after Virus Challenge | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group | Animal ID | 0 | 1–9 | 10–15 | 16–20 | 21–25 | 26–30 | 31–40 | D41–D50 |
| H-G1 | Hare #231 Not vac | Virus challenge with 1 mL (100 ffu/mL) of ha-MYXV (38455PT18) | N | D11-very mild eyelid oedema. <25% of eye closure. D13—mild eyelid oedema. <25% of eye closure. D14—moderate eyelid oedema. 50% of eye closure. | D16 marked eyelid oedema. 50–75% of eye closure. | D25 marked eyelid oedema. >75% of eye closure. | D30-marked eyelid and anogenital oedema. 100% of eye closure. Anorexia and dyspnea. Euthanasia. | ||
| Hare #232 Not vac | N | D12—mild eyelid oedema. <25% of eye closure. D14—moderate eyelid oedema. 25–50% of eye closure. | D18 marked eyelid oedema. 50–75% of eye closure. | D25 marked eyelid oedema. >75% of eye closure. | D30 marked eyelid and anogenital oedema. 100% of eye closure. Anorexia and dyspnea. Euthanasia. | ||||
| Hare #233 Not vac | N | N | N | N | D30—Second virus challenge with 1000 ffu of ha-MXYV (38455PT18), day 102 in Figure 1. | D40—very mild eyelid oedema. <25% of eye closure. D42—mild eyelid oedema. <25% of eye closure. | D45—Marked eyelid, foreskin and anogenital oedema. 50–75% of eye closure. Anorexia. D50-Death | ||
| H-G2 | Hare #076 1× M-FSA 1× M-H | N | D16—very mild eyelid oedema. <25% of eye closure. D18—very mild eyelid oedema. <25% of eye closure. | D20—mild eyelid oedema. <25% of eye closure. | D20—moderate eyelid oedema. 25–50% of eye closure. D30—marked eyelid oedema. 50–75% of eye closure. | D40—marked eyelid oedema. >75% of eye closure. | D48—marked eyelid and foreskin oedema. >75% of eye closure. Anorexia and dyspnoea. Euthanasia. | ||
| #078 Hare 1× M-FSA 1× M-H | N | D12—very mild eyelid oedema. <25% of eye closure. | D16—mild eyelid oedema. <25% of eye closure. | D20—mild eyelid oedema. <25% of eye closure. Death | |||||
| H-G3 | Hare #042 10× M-FSA 10× M-H | N | N | N | N | D30—Second virus challenge with 1000 ffu of ha-MXYV (38455PT18), day 102 in Figure 1. | D40—very mild eyelid oedema. <25% of eye closure. D45—mild eyelid oedema. <25% of eye closure. | D50—mild eyelid oedema. <25% of eye closure D60—moderate eyelid, testis and foreskin oedema. 25–50% of eye closure. Anorexia. Death. | |
| Hare #043 10× M-FSA 10× M-H | N | D12-very mild eyelid oedema. <25% of eye closure. | D16—Left l ower eyelid erythema. | D25—N | N | N | N | ||
| Hare #044 10× M-FSA 10× M-H | D7—mild eyelid oedema. <25% of eye closure. | D12—mild eyelid oedema. <25% of eye closure. | D14-D20—moderate eyelid oedema. 25–50% of eye closure. | D21—Beginning of crusting and improvement of eye opening. | D26—N, in addition to scars on the eyelids. | ||||
| H-G4 | Hare #10 Natural immunity | N | N | N | N | D30-Second virus challenge with 1000 ffu of 38455PT18. | N | N | |
| Days after Virus Challenge | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Group | Animal ID | 0 | 1–9 | 10–15 | 16–20 | 21–25 | 26–30 | 31–40 | D41–D50 |
| RG-1 Not vac | Rabbit #000 | Virus challenge of 1000 ffu of ha-MYXV (38455PT18), day 65 in Figure 1. | D4—very mild eyelid oedema. No genital changes. D8—moderate eyelid and genital oedema. 50–75% of eye closure. | D11—Marked eyelid, foreskin and genital oedema. >75% of eye closure Anorexia. Death. | |||||
| Rabbit #001 | D5—very mild eyelid and genital oedema. D8—moderate eyelid and genital oedema. 50–75% of eye closure. | D11—Marked eyelid, foreskin and genital oedema. >75% of eye closure Anorexia. Death. | |||||||
| Rabbit #449 | Virus challenge of 1000 ffu of ha-MYXV (20545PT20), day 35 in Figure 1. | D8—very mild eyelid oedema. <25% of eye closure. No genital changes. | D12—Very mild eyelid and foreskin oedema. <25% of eye closure. No genital oedema. Death. | ||||||
| Rabbit #451 | D8—very mild eyelid oedema. <25% of eye closure. No genital changes. | D12—Very mild eyelid and foreskin oedema. <25% of eye closure. No genital oedema Death. | |||||||
| RG-2 1× M-H | Rabbit #442 | N | N | N | N | N | N | N | |
| Rabbit #444 | N | N | N | N | N | N | N | ||
| Rabbit #445 | N | N | N | N | N | N | N | ||
| RG-3 1× M-RHD | Rabbit #446 | N | N | N | N | N | N | N | |
| Rabbit #447 | N | N | N | N | N | N | N | ||
| Rabbit #448 | N | N | N | N | N | N | N | ||
| Group | Animal ID | Challenge | Pathological Examination | Bacteriological Examination from Liver, Spleen and Lungs | Parasitological Examination | ||||
|---|---|---|---|---|---|---|---|---|---|
| Eyelid/Ano-Genitalia/Lip | Liver | Spleen | Lungs | Kidney/Other | |||||
| H-G1 | Hare #231 | 100 ffu of ha-MYXV isolated from hare (38455PT18) | Macroscopic: Marked eyelid, lips, genitalia and anus oedema. Thickening and congestion of the scrotal sac wall with necrosis. Accumulation of fibrin within the scrotal pouches. Congestion of the testicles. Microscopic: Eyelid with necrosis of the epidermis and conjunctiva with bacterial infiltration. Scrotum with oedema at the dermo-epidermal junction with detachment of the epidermis. Lip with necropurulent dermatitis and presence of extensive bacterial clusters. Testis with absence of germ cells, persisting only Sertoli cells. Epididymis with necrosis of the lining epithelium and accumulation of necrotic cells in the lumen of the ducts. Vas deferens with accumulation of myxoid tissue underlying the lamina propria. | Moderate periportal infiltration by mononuclear cells | N | N | Kidney with perivascular lymphocytic infiltration. Empty stomach. | Escherichia coli and Stenotrophomonas maltophilia | Mild infection by Eimeria spp. |
| Hare #232 | Macroscopic: Marked eyelid, lips, genitalia and anus oedema. Microscopic: Eyelid with epidermal hyperplasia and extensive proliferation of myxoid tissue in depth to the conjunctiva. Lips with epidermal hyperplasia, ballooning degeneration of keratinocytes, intense production of myxoid tissue, infiltration of muscle tissue by mononuclear cells and heterophils. Prepuce with extensive and severe necropurulent dermatitis with strong bacterial infiltration, myxoid tissue in the dermis, diffuse infiltration by heterophils in depth, necropurulent foci. | Cellular infiltration, namely by heterophils in the porta spaces and occasionally in the sinusoid capillaries. | Marked depletion of lymphocytes. | Multifocal purulent alveolitis. | Kidney with perivascular infiltration by mononuclear cells. Empty stomach. | Stenotrophomonas maltophilia, Staphylococcus equorum and Staphylococcus xylosus | Mild infection by Eimeria spp. | ||
| Hare #233 | 1000 ffu of ha-MYXV isolated from hare (38455PT18) | Macroscopic: Nodular thickening on the eyelids and lips and oedema of the prepuce. Microscopic: Eyelid with epidermal hyperplasia and vacuolization of keratinocytes. Proliferation of myxoid tissue in the dermis. Diffuse microhaemorrhages and foci of infiltration by heterophils. Conjunctival epithelial hyperplasia and underlying stromal oedema. Lips with epithelial hyperplasia and marked vacuolization of keratinocytes with proliferation of myxoid tissue in the dermis and intense infiltration by heterophils. Foreskin with thick necropurulent exudate and extensive haemorrhages, presence of myxoid tissue and infiltration by heterophils. | N | Marked depletion of lymphocytes. | Lung collapse. | Empty stomach. | Escherichia coli and Enterobacter sakazakii | Mild infection by Eimeria leporis. | |
| H-G2 | Hare #076 | 100 ffu of ha-MYXV isolated from hare (38455PT18) | Macroscopic: Marked oedema of the prepuce, eyelids and lips. Empty stomach. Microscopic: Eyelid with dermal oedema. Lips with foci of epithelial hyperplasia and vacuolization of keratinocytes. Foreskin with marked oedema of the dermis. | Liver with periportal infiltration by mononuclear cells and heterophils in sinusoid capillaries. | Mild depletion of lymphocytes. | Congestion. | Escherichia coli | N | |
| Hare #078 | Macroscopic: Marked oedema of the foreskin, eyelids and lips. Presence of small ulcers on the lips. Microscopic: Lips with foci of epithelial hyperplasia and vacuolization of keratinocytes. Foreskin with marked oedema of the dermis. Epididymis with oedema and extensive haemorrhages in the basement membrane zone. Hyperplasia of the duct lining epithelium and desquamation of the ductal epithelium. Infiltration by heterophils. Eyelid with dermal oedema. | Liver with periportal infiltration, mainly by mononuclear cells and heterophils in sinusoid capillaries. | Marked depletion of lymphocytes. | Congestion. | Kidney with the presence, mostly perivascular, of small clusters of lymphoid cells. Empty stomach. | Enterococcus gallinarum | Marked infection by Eimeria spp. | ||
| H-G3 | Hare #042 | 1000 ffu of ha-MYXV isolated from hare (38455PT18) | Macroscopic: Nodular thickening and moderate oedema and nodular thickening of eyelids, nose, lips and testis. Oedema of the extremities of the hind limbs. Microscopic: Eyelid with epidermal hyperplasia and vacuolization of keratinocytes. Moderate proliferation of myxoid tissue in the dermis. Lips with necropurulent lesions, moderate epithelial hyperplasia and vacuolization of keratinocytes with proliferation of myxoid tissue in the dermis and infiltration by heterophils. Foreskin with moderate presence of myxoid tissue and infiltration by heterophils. | Congestion. | Marked depletion of lymphocytes. | Congestion. | Empty stomach. | Vibrio vulnificus | N |
| Hare #044 | 100 ffu of ha-MYXV isolated from hare (38455PT18) | Macroscopic: Mild scar on the eyelids. | N | N | N | N | N | N | |
| R-G1 | Rabbit #449 and Rabbit #451 | 1000 ffu of ha-MYXV isolated from rabbit (20545PT20) | Macroscopic: Very mild oedema of the eyelids and lips. Presence of extensive pleural effusion with fibrin. Microscopic: Eyelid with mild oedema of the deep dermis. | Congestion of Liver (#449). | Spleen with total necrosis of lymphoid follicles with only a very congested red pulp visible. | Lung congestion and oedema. Pleural efusion. Necrosis of peribronchial lymphoid tissue. | Full stomach. | N | N |
| Rabbit #000 and Rabbit #001 | 1000 ffu of ha-MYXV isolated from hare (38455PT18) | Macroscopic: Marked swelling of the eyelids, lips and external genitalia. Presence of purulent mucous exudate on the eyelids. Reduced content in the stomach. Microscopic: Eyelid with marked oedema, small foci of myxoid tissue in the dermis, purulent conjunctivitis of bacterial etiology. Lip with oedema and small foci of myxoid tissue. | N | Spleen with congestion and necrosis of lymphoid follicles. | N | N | N | Medium infection with Eimeria perforans. | |
| ID | Vaccination Data | Viral Loads in Different Tissues (DNA Copies/mg Tissue) | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1st | 2nd | Liver and Spleen | Lung | Duodedum | Kidney | Eyelid | Lip | Genitalia | Urine | Seminal Vesicle | Faeces | Brain | Bone Marrow | Spinal Cord | |
| Hares | |||||||||||||||
| #231 | Not vac | Not vac | 1.60 × 107 | 1.47 × 108 | 5.76 × 106 | 1.71 × 107 | 2.88 × 109 | 1.75 × 109 | 2.23 × 107 | 7.44 × 106 | 1.78 × 109 | 3.98 × 108 | 4.05 × 106 | 1.31 × 106 | 1.53 × 106 |
| #232 | Not vac | Not vac | 1.10 × 108 | 1.23 × 108 | 3.66 × 107 | 3.15 × 109 | 1.24 × 1010 | 7.47 × 109 | 1.24 × 1010 | 5.47 × 103 | 5.42 × 106 | 1.83 × 106 | 4.95 × 105 | 3.70 × 105 | 5.45 × 106 |
| #233 | Not vac | Not vac | 6.73 × 108 | 6.28 × 108 | 4.03 × 106 | 4.39 × 108 | 1.34 × 1010 | 3.88 × 109 | 5.33 × 1010 | NT | 5.37 × 105 | 1.52 × 108 | 2.29 × 108 | 6.83 × 104 | 9.91 × 104 |
| #076 | 1× M-FSA | 1× M-H | 2.55 × 105 | 2.33 × 105 | 5.59 × 104 | 1.85 × 106 | 1.34 × 107 | 1.11 × 106 | 2.08 × 108 | 2.16 × 105 | ND | 1.16 × 105 | ND | 3.11 × 103 | ND |
| #077 | 1× M-FSA | 1× M-H | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| This animal was not submitted to challenge but was analysed to MYXV to eliminate as the cause of death | |||||||||||||||
| #078 | 1× M-FSA | 1× M-H | 1.17 × 108 | 4.39 × 108 | 7.20 × 107 | 2.22 × 108 | 2.89 × 1010 | 4.16 × 1010 | 5.40 × 109 | NT | 1.36 × 105 | 1.50 × 109 | 6.26 × 106 | 3.22 × 104 | 8.54 × 106 |
| #042 | 10× M-FSA | 10× M-H | 2.90 × 107 | 3.06 × 107 | 8.72 × 106 | 1.50 × 107 | 8.46 × 109 | 1.08 × 109 | 7.73 × 109 | NT | NT | 7.51 × 108 | 7.42 × 104 | 9.25 × 104 | 2.36 × 105 |
| #043 | 10× M-FSA | 10× M-H | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| #044 | 10× M-FSA | 10× M-H | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND |
| #10 | Natural immunity | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| Rabbits | |||||||||||||||
| #442 | 1× M-H | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| #444 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ||
| #445 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ||
| #446 | 1× M-RHD | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | |
| #447 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ||
| #448 | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ND | ||
| #449 | Not vac | 2.30 × 1010 | 2.03 × 1010 | 7.16 × 108 | 9.97 × 108 | 1.29 × 1010 | 1.36 × 109 | 4.19 × 1010 | 2.84 × 107 | ND | 2.22 × 104 | ND | 1.02 × 102 | ND | |
| #451 | 1.24 × 1010 | 3.48 × 1010 | 1.04 × 108 | 4.94 × 109 | 1.87 × 1010 | 9.97 × 108 | 3.01 × 109 | 8.72 × 106 | NT | 1.15 × 104 | ND | 8.91 × 102 | ND | ||
| #000 | Not vac | 9.91 × 109 | 7.74 × 1010 | 1.25 × 108 | 1.45 × 109 | 6.98 × 1010 | 2.33 × 109 | 6.07 × 109 | 1.15 × 108 | NT | 1.24 × 105 | 3.13 × 104 | ND | ND | |
| #001 | 3.32 × 108 | 2.11 × 109 | 2.93 × 107 | 8.32 × 109 | 9.02 × 1010 | 1.89 × 108 | 5.04 × 1010 | 2.04 × 106 | NT | 2.25 × 103 | 4.12 × 103 | 2.15 × 100 | ND | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abade dos Santos, F.A.; Carvalho, C.L.; Valente, P.C.L.G.; Armés, H.; Reemers, S.S.; Peleteiro, M.C.; Calonge Sanz, I.; Dalton, K.P.; Parra, F.; Duarte, M.D. Evaluation of Commercial Myxomatosis Vaccines against Recombinant Myxoma Virus (ha-MYXV) in Iberian Hare and Wild Rabbit. Vaccines 2022, 10, 356. https://doi.org/10.3390/vaccines10030356
Abade dos Santos FA, Carvalho CL, Valente PCLG, Armés H, Reemers SS, Peleteiro MC, Calonge Sanz I, Dalton KP, Parra F, Duarte MD. Evaluation of Commercial Myxomatosis Vaccines against Recombinant Myxoma Virus (ha-MYXV) in Iberian Hare and Wild Rabbit. Vaccines. 2022; 10(3):356. https://doi.org/10.3390/vaccines10030356
Chicago/Turabian StyleAbade dos Santos, Fábio A., Carina L. Carvalho, Pâmela C. L. G. Valente, Henrique Armés, Sylvia S. Reemers, Maria C. Peleteiro, Inés Calonge Sanz, Kevin P. Dalton, Francisco Parra, and Margarida D. Duarte. 2022. "Evaluation of Commercial Myxomatosis Vaccines against Recombinant Myxoma Virus (ha-MYXV) in Iberian Hare and Wild Rabbit" Vaccines 10, no. 3: 356. https://doi.org/10.3390/vaccines10030356