
Enhancing Carbon Sequestration in Mediterranean Agroforestry Systems: A Review


1
INIAV-Instituto Nacional de Investigação Agrária e Veterinária, GREEN-IT, ITQB NOVA-Instituto de Tecnologia Química e Biológica/NOVA Universidade de Lisboa, Nova Oeiras, 2784-505 Oeiras, Portugal
2
INIAV-Instituto Nacional de Investigação Agrária e Veterinária, 1300-596 Lisboa, Portugal
3
CEF-Centro de Estudos Florestais, Instituto Superior de Agronomia, Universidade de Lisboa, 1349-017 Lisboa, Portugal
*
Author to whom correspondence should be addressed.
Agriculture 2022, 12(10), 1598; https://doi.org/10.3390/agriculture12101598
Submission received: 27 August 2022
/
Revised: 12 September 2022
/
Accepted: 27 September 2022
/
Published: 2 October 2022
(This article belongs to the Special Issue Soil Organic Matter and Its Role in Soil Fertility)
Abstract
:The agroforestry systems with a high potential for C sequestration are those degraded by poor management strategies. Studies on changes in soil C status in these ecosystems mostly take into account labile C pools. Labile and stable soil organic matter (SOM) fractions are affected by soil management and land-use changes. Stable C pools are essential to understanding effects of land-use on soil C storage in the long term. The SOM stability is partly enhanced by the interaction of SOM with minerals and its inclusion into soil aggregates. Recalcitrant substances (e.g., lignin and chitin) also contribute to the passive SOM fraction. Macroaggregates mostly reflect the influence of plant roots and coarse intra-aggregate particulate SOM (POM), whereas microaggregates reflect the influence of fine interaggregate POM, clay concentration and humified SOM fraction. Often, POM is more sensitive to soil management changes than total SOM. Glomalin is a recalcitrant protein consisting of chitin produced by mycorrhizal fungi to protect hyphae. Glomalin has implications on C sequestration in agroforestry soils, but further research is needed before any prediction can be made. One challenge is reducing the CO2 emission from roots, and increasing the recalcitrant root C.
1. Introduction
Anthropogenic activities have adversely affected the global carbon (C) cycle and contributed to global climate change. A major driver of recent changes in the earth’s climate is the increase in atmospheric carbon dioxide (CO2). The amount of soil organic C at surface (~1500 Gt C) stored in soil organic matter (SOM) as decomposed plant litter and residues [1] is about three times greater than that of C in the atmosphere. Therefore, the accumulation of C in soil can partially offset the effect of anthropogenic CO2 emission and is an important strategy to mitigate greenhouse gases (GHG) [2]. The major factors influencing soil C storage are environmental (temperature, precipitation, and soil) and biotic (plant species and composition and anthropogenic activities).
The land use systems were ranked in terms of their soil organic carbon (SOC) concentration by Nair et al. [3] in the order: forest > agroforestry > other tree plantation (e.g., orchards) > arable crops. The transition from an agricultural to an agroforestry system is beneficial to SOC; in contrast, the conversion of secondary and primary forests to agroforestry systems leads to SOC losses [4]. The large differences in C sequestration in soils among land uses reflect the biophysical and socioeconomical characteristics of the system parameters and/or the methodology used. The agroforestry system was recognized by the land use, land use forestry change (LULUCF) as a C sequestration activity, especially under afforestation and reforestation activities. IPCC [5] estimated that 630 million ha of unproductive croplands could be converted to agroforestry worldwide, with the potential to sequester 586,000 t C yr−1 by 2040. However, Nair [6] pointed out that there is a significant lack of rigorous data in C sequestration in these systems. To assess changes in SOC stock and storage/loss following a land use change (LUC), some basic data should be considered such as the description of previous land use, SOC stocks, or concentrations and bulk densities in both the previous and new land use, soil depth considered, and time span since conversion [6,7]. The loss of SOC following LUC can be very fast, while usually, it takes a long time to gain. The main difficulty to properly assess SOC changes in agroforestry systems compared to other land uses is associated with the spatial heterogeneity caused by scattered trees and the cultivation of different plant species, which induce different gradients in organic inputs to the soil, and a large number of soil samples are necessary to explore such heterogeneity. Only a few studies have measured the SOC storage in deep soil layers of agroforestry systems [4].
In Europe, and especially in the Mediterranean basin, the abandonment and transformation of large areas of cropland into pastures in recent decades, including agroforestry systems, has been favored with the aim of reducing the surplus of agriculture production and enhancing the C sink capacity of the soils. After the land abandonment, secondary succession vegetation started with pasture colonization affecting strongly the SOC [7,8]. The permanent pastures in agroforestry systems could store 50–60 t C ha−1 [9].
However, soils have a limited potential to sequester C (sink saturation) when the annual benefits reach a quasi-equilibrium (steady state) [10,11]. In fact, Dungait et al. [11] reported that it is unlikely that all ancient C in soils can be physically protected because there must be a threshold at which the available C storage niches are filled, referred to as C saturation and defined by the physicochemical properties of the soil. Six et al. [10] reinforced the need for the mechanistic explanation of a saturation level for the unprotected and biochemical protected pools as a priority. In order to account for this, the IPCC [12] recommended at least a 20-year period for soil C sequestration to account for the national GHG inventories.
Within this context, the specific objectives of the present review were: (1) to review the concepts for C sequestration in soil, understanding the terminology and the context in which each concept is used; (2) to discuss the land use strategies under Mediterranean conditions to enhance C sinks by the contribution of agroforestry systems; (3) to evaluate the potential contribution of belowground plant tissue to C stock in agroforestry soils; and (4) to address new approaches for the appropriate increasing of C sequestration in Mediterranean agroforestry pasture soils.
2. Concepts for Carbon (C) Sequestration in Soil and Understanding the Terminology
Very often, the term “C sequestration” in soil is misunderstood and is used to refer the annual accumulated C in soil (stock). Ideally, the latter should be reported as rate (mass per unit of area and time). Accumulated soil C includes the stable OM pool and the labile (active) OM fraction, with different residence times. The labile fraction is generally more sensitive to soil management changes than the stable OM (passive and recalcitrant), and is then more sensitive than the total SOM [8,13,14]. The labile fraction is constituted by partially decomposed residues and represents 20–50% of the total SOM in cold and semiarid regions [15], and only 10–20% of SOM in tropical and subtropical regions [15,16]. Therefore, the active pool is that which would change mostly in a 1–10-year period. Batjes [17] reported that the most rapid changes in SOM concentration occurred during the first five years after a management practice was imposed, with slower changes occurring thereafter. The passive (recalcitrant) and slow-release OM pools have meant residence times of hundreds to thousands of years [3,18] and are referred as sequestered C in soil.
Based on C dynamics and homogeneity in terms of residence times, SOM may be divided into four pools (Table 1): an “active” or “labile” pool of readily oxidized compounds coming from fresh plant material and microbial biomass; a “slowly oxidized pool” (moderate pool) associated with macroaggregates, depending on soil mineralogy and aggregation, as well as agronomic practices; a “very slowly oxidized pool (slow pool)” associated with microaggregates, where the main controlling factor is the water stability of aggregates (agricultural practices have only small effects); and a “passive or recalcitrant pool” where clay mineralogy is the main protecting factor for microbial attack (cultural practices have no relevant effects) [11,13,14,17].
The physical protection of SOM includes its occlusion within aggregates and the adsorption onto minerals [11]. The term “humic substances” (humic and fulvic acids, humin) is used to characterize chemically the SOM, humic acid, and humin fractions being considered stabilized/passive and inaccessible to soil microorganism forms [11,19]. Humic substances have been contested by some authors. For instance, Lehmann and Kleber [20] suggested the abandonment of terms related with humic substances because they consider those terms referring to old analytical methods, based on alkaline extraction of SOM fractions. However, Ponge [21] proposed keeping them considering their importance in measuring soil ecosystem services. Therefore, this question is still open. In terms of soil C sequestration, the stability of stored SOM is more important than its quantity. Sequestered C is enhanced when it is in forms physically (e.g., inside the stable microaggregates, forming organo–mineral complexes as secondary particles; this is particularly relevant when 2:1 clay mineral dominates in soil) or chemically protected from microbial activity, or stored in recalcitrant compounds (chitin, cutin, and suberin) with a long turnover, i.e., a slow rate of decomposition [10,11,22,23].
Gabarrón-Galeote et al. [8] described the nonprotected SOC easily decomposable (free POM), with a short turnover time (1–10 years), the physically protected SOC pool by occlusion in stable microaggregates, with an intermediate turnover time (10–100 years); the chemically protected SOC pool by organo–mineral association with silt and clay particles, also with a turnover time of 10–100 years; and a biochemically recalcitrant SOC pool with a turnover time over 100 years. These authors [8] emphasized that knowing the degree of protection of the SOC, expressed as SOC fraction, is crucial for a better understanding of SOC dynamics in soil.
The most basic question is about the C stock increase because there is already a large amount of C stored in soils. Since it is hard to measure a small variation in a large C pool (total SOC) with a high level of precision, an alternative approach would be to select a specific SOC pool within the total SOC as an indicator of change in C stock. Procedures that measure specific physical or chemical labile fractions (i.e., the particulate OM-POM, microbial biomass, and other pools) are useful as indicators of land use changes and for the development of models, but they are complex and time-consuming and would not be run on as many samples as total soil C [18]. Wander et al. [24] reported a greater increment of POM (100%) in the top 5 cm layer of no-tilled silty loam soils than the total SOC (more than 30%).
Consequently, total C measured by the dry combustion (loss in ignition method) or the wet oxidation method [25] is not appropriate to assess components and dynamics regarding soil C stock in the long-term issue (C sequestration). Besides being less reliable, these methods are also time-consuming and more dangerous for the environment, and do not discriminate the labile from that corresponding to humic substances and other recalcitrant organic compounds (nonlabile forms). The innovative, nondestructive techniques such as nuclear magnetic resonance (NMR) spectroscopy and inelastic neutron scattering (INS) (technique under development) have a high potential for SOC monitoring in the field. The NMR distinguishes structures that are characteristics of newly formed SOM, in particular the POM in the sand fraction [26,27,28]. This issue remains of critical importance for the C sequestration debate.
Several variations in physical (particle size, aggregate size, or density) and chemical fractionation, or a combination of both, have been employed to separate the more actively cycling (labile SOM) from more stable (recalcitrant SOM) [23]. Because these methods are operational, not functional, none of them perfectly isolate a single active fraction [23]. Both pools can be separated by size and/or density separation. The density fractionation method uses a high-density solution to isolate SOM fractions that differ in stability (separation from soil matrix). It separates soil C into light (>150 μm) and heavy fractions (LF and HF, respectively). The LF is primarily composed of younger plant debris, only partially decomposed and still resembling their original material, but also contains seeds and microbial debris, such as fungal hyphal and spores which are good indicators of labile fraction [10], and including the POM. The LF is controlled by soil management and may be used as an early indicator of effects of soil management changes on SOM [23,29,30]. The HF fraction is composed of amorphous material in an advanced stage of humification associated with soil minerals, forming organo–mineral complexes that are thought to be largely recalcitrant C. Therefore, HF is more crucial to soil C sequestration compared with LF [29,31].
By physical fractionation, particle-size fractions are distributed as coarse sand, fine sand, silt, clay, and particulate organic matter (from the coarse-sand-sized fraction (POM-CS) and fine sand (POM-FS)) [32]. Soil organic matter in the sand or coarser fractions consist of relatively unaltered organic material (POM) [29]. Furthermore, the coarse intra-aggregate POM is physically fragmented and chemically decomposed into fine intra-aggregate POM (in the particle fraction of 50/53–250 µm). This intermediate OM fraction contains fine sand, fine free SOM easily decomposable (POM), stable microaggregates that contain physically stabilized SOC, and biochemically recalcitrant SOM [8,16]. The fine intra-aggregate POM gradually becomes encrusted with silt and clay particles and bound by microbial products (e.g., glomalin, excreted by soil arbuscular mycorrhizal fungi) and is chemically stabilized forming very stable microaggregates (<50/53 µm) within the macroaggregates (<2 mm). The biochemical/protected recalcitrant SOM is crucial for the storage and stabilization of SOC in the long term [8,10,11,22,33,34], contributing to the improvement in soil structure. The physical protection is like a first step in the conservation of SOM that slows down the mineralization process through isolation of microbes from their organic substrates and/or water and O2 supplies [10,22,35,36]. The size of soil aggregates is usually inversely proportional to the amount of energy needed to disrupt them; then, the microaggregates afford the most protection to the associated SOM [22]. By soil disturbance, like the cultivation practice, the macro- and microaggregates’ stabilization can stop, resulting in lower C sequestration in soil [10,22,34]. Then, the loss of SOM is a key factor decreasing the aggregate stability.
Particulate organic matter (POM) is often reported as a stable SOM fraction, but as previously reported, it is not sufficiently stable to explain the long-term changes in total SOM [8,10,13].
Nunes et al. [37] used a meta-analysis for soil management assessment framework (SMAF) as a comprehensive tool to assess the effects of land use and management practices on soil health by integrating multifunctional indicators (e.g., SOC, aggregate stability, microbial C, pH, P, and bulk density) which represent the important soil functions. They confirmed that tillage generally decreased SOC content by disrupting soil aggregates, by increasing mineralization rates (i.e., oxidation of C), and by exposing bare soils to erosion from water and wind. In addition, the long-term loss of SOC led to reduced soil biological activity, aggregate stability, water storage, and incrusting surface crusting and erosion. They advertised that conversion of conventional tillage to no-tillage improved biological and physical indicators of soil health in the topsoil, likely due to the maintenance of crop residues on the soil surface, and a subsequent increase in SOM. Finally, they recommended that converting annual cropping systems to perennial systems is a strategy to enhance long-term agroecosystem sustainability, environmental protection, and climate change resilience.
Actual measurements to understand SOM pool sizes, and how they will change with management systems, might be of great relevance and interest for policymakers and traders of C credits. Such measurements can be taken by modeling (e.g., the Q model, particularly advantageous under changing climatic conditions; the Roth C model, where “DPM” represents the decomposing plant material and “BIO” represents the biomass pools and simulates the physical protection in the long term; and the Century model, where the metabolic compounds are associated with the “active” and “metabolic” pools) [11]. However, modeling can only be achieved if there is a large number of on-site measurements at specific sites to verify the model. Soil sampling and analytical methods require further studies. The nondestructive techniques under development would probably be of high importance for this purpose.
3. Land Use Strategies under Mediterranean Conditions to Enhance Carbon (C) Sinks: Contribution of Agroforestry Systems
Perennial crops and management practices such as no-tillage, residue mulching, extended crop rotation, and cover cropping are strategies that may contribute to the soil C stock in Mediterranean areas, contributing to reducing the agricultural GHG emission. Combining some of these characteristics, agroforestry systems have a great potential for mitigation of CO2 by C sequestration. However, in order to realize the full potential of these systems for climate change mitigation, other GHGs such as methane (CH4) and nitrous oxide (N2O) should be taken into account because there is a lack of studies on GHG emissions from agroforestry systems to fully support that.
Agroforestry systems are complex agroecosystems combining trees (mostly evergreen trees) and crops (e.g., cereals), or trees and permanent pastures (native or improved), mostly permanent, within the same field [4,38]. These systems include a wide variety of tree and crop species arrangements according to the climatic region. Besides the C sequestration, they can also provide other ecosystem services such as an improvement in soil fertility, protection of soil erosion, enhancement of biodiversity, and the climate change regulation. These cropping systems have attracted particular attention as a C sequestration strategy from both industrialized and developing countries. In industrialized countries, farmers are compensated for C sequestration in the agroforestry systems based on the amount of C sequestered and the market price, whereas in developing countries, farmers benefit from any gain in productivity of these ecosystems associated with the adoption of agricultural practices to improve C sequestration [3].
In agroforestry, trees, shrubs, and permanent pastures can mitigate the atmospheric CO2. The C storage in these soils implies the removal of CO2 from the atmosphere through crop photosynthesis (C capture) and deposition in sinks (C storage) in the soil as a stable OM pool. The capacity of soils and biomass in these systems to store C depends on several factors, including environmental factors, land use, tree density, plant species and age, and management practices [4,7]. Most results showed that SOC in deeper layers and the relatively stable soil C were derived from tree components (C3 plants), and, therefore, the tree-based pasture system has a greater potential for C sequestration as compared with the treeless system (pasture/grassland) [3].
Among the agroforestry systems in the Iberian Peninsula, in Southern Europe, the “montado” in Portugal and “dehesa” in Spain are of great relevance. They consist mostly of multipurpose trees, such as chestnuts (Castanea spp.), oaks (Quercus spp.), and olives (Olea europa L.) with understory vegetation such as pastures with grazing from extensive livestock (sheep, black pigs, and cattle), usually in a rotation scheme, or the recurrent cereal cropping. This practice is old in Europe, in particular in the Mediterranean basin, and occupies about 4 million hectares in Portugal and Central and SW Spain [7,38,39].
The Mediterranean agroforestry systems generally have low soil fertility and are limited by their shallow soil depth and low water and nutrient availability, resulting in natural pastures with poor productivity and quality [38]. After noticing the low productivity of pastures, farmers in the Iberian Peninsula began to sow legume-rich mixture pastures [40] (Figure 1), which contributed to a simultaneous increase in pasture productivity and SOC concentration, and prevented soil degradation.
Although improved pastures can stay highly productive for longer than 20 years under proper grazing and fertilization, no studies are found to clearly indicate the recommended pasture management relating to their persistence.
Together with the trees, permanent pastures (natural or improved) also provide a fast way to build soil C for several reasons: firstly, the presence of perennial species which grow continuously rather than seasonally with extensive root systems; secondly, the minimal soil disturbance relative to arable crops or annual pastures; thirdly, the facilitation of soil organisms’ diversity and activity, which promote soil aggregation and therefore C protection; and finally, the avoidance of erosion due to the increment of soil cover, depending on pasture management [41]. Managing the degraded pastures has a great potential for C sequestration, but it is difficult to quantify this potential in the field because it requires studying the stable and mature ecosystems.
Evaluation of spatial heterogeneity of soil C in these agroforestry systems has been carried out mainly at tree level, showing that scattered trees enhance soil fertility and improve soil physical and biological condition beneath their canopies [40,41]. Gómez-Rey et al. [42,43] measured a significantly higher amount of organic C in the 0–10 cm soil layer beneath the tree canopy in both unmanaged (2.4 kg C m−2) and improved (3.1 kg C m−2) Spanish pastures, as compared with the open field (1.8 and 2.1 kg C m−2, respectively, in natural and improved pastures). The increments of SOC concentration in the surface layer beneath the tree canopy suggested a significant relationship with the amount of organic litterfall residues annually deposited on the soil surface [44], and with the root litter produced in the topsoil layer [45]. This variability decreased with the increase in distance to the tree trunk [39,42]. Indeed, Simón et al. [39] found a positive correlation between the presence of trees and SOC stocks up to an 8 m distance from their crowns. In improved pastures, Gómez-Rey et al. [43] found that SOC concentration was greatly affected by tree presence, with higher content in soil beneath the tree canopy than in the open field, while in unmanaged pastures, the C/N ratio values were of the same magnitude in both sites in the litterfall. Tree litterfall residues are in general slow to decompose in comparison with the herbaceous vegetation due to the higher C/N ratio. Gomez-Rey et al. [43] verified that this organic C corresponded mostly (58–79%) to the nonparticulate OM fraction. The belowground plant material could also contribute to the nonparticulate SOM but no data are available. Pulido-Fernández et al. [46] found 2.3% SOC in the top 5 cm soil layer of woodlands of SW Iberia, with a large soil variability. SOC was relatively high beneath the tree canopy (average 3.4%) and less concentrated and reduced in variability with depth. In Central and Western Spain, Howllet et al. [47] also observed that the scattered trees in the agroforestry systems had a strong influence on spatial variability in SOC concentration, with a greater soil C storage underneath tree canopies. According to Simón et al. [39], montados/dehesas have scarcely been included in large-scale studies to assess SOC sequestration capacity. This is probably due to spatial heterogeneity, complex management, and their generally low SOC contents. Pulido-Fernández et al. [46] found 2.3% of organic C in the top 5 cm soil layer of woodlands of SW Iberia, with a large soil variability. SOC was relatively high beneath the tree canopy (average 3.4%) and less concentrated and reduced in variability with depth. In Central and Western Spain, Howllet et al. [47] also observed that the scattered trees in the agroforestry systems had a strong influence on spatial variability of organic C concentration, with a greater soil C storage underneath tree canopies. According to Simón et al. [39], montados/dehesas have scarcely been included in large-scale studies to assess SOC sequestration capacity. This is probably due to spatial heterogeneity, complex management, and their generally low SOC contents.
It is noteworthy that the effect of scattered trees on the accumulation of organic C in the topsoil might be enhanced by the installation of improved pastures. Indeed, Gómez-Rey et al. [42] reported that the increase in the soil organic C stock up to a 20 cm depth, in a 26-year-old improved pasture, was 0.18 and 0.84 kg m−2, respectively, in open and tree-covered areas, as compared to a natural pasture with shrub control every six years.
Trees in oak woodlands may create heterogeneity regarding the C/N ratio in the topsoil, depending on the nature of the understory. In fact, with improved pastures, Gómez-Rey et al. [42] reported a higher C/N beneath a tree canopy than in the open, while in unmanaged pastures, the C/N ratio values were of the same magnitude in both sites. This trend agrees with Rodrigues et al. [48], who observed a decrease in the C/N ratio from 33, in areas with trees and shrub understory, to 20 in the same area with improved pastures. Such a difference may be associated with tree litterfall residues which, in general, are slow to decompose in comparison with herbaceous vegetation due to the higher C/N ratio. In addition, Gómez-Rey et al. [43] verified that the organic C beneath trees corresponded mostly (58–79%) to the nonparticulate SOM fraction. The belowground plant material could also contribute to the nonparticulate SOM but no data are available.
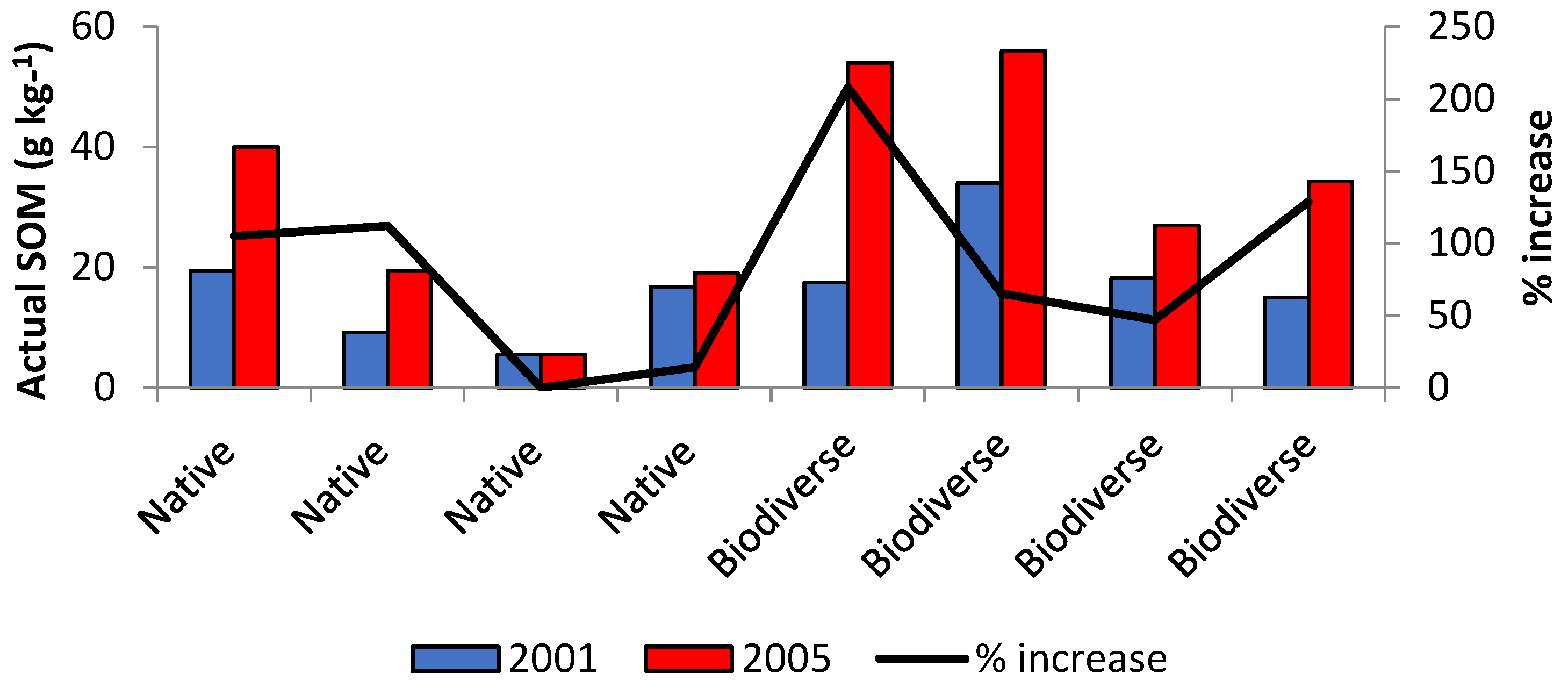
The environmental protection and monetary return are the main drivers for agroforestry in industrialized nations. Since 2009, the Portuguese government have paid farmers through the Portuguese Carbon Fund [49] to improve degraded pastures, in a total area of 42,000 ha, with the aim of accumulating EUR 0.91 million of CO2eq from 2010 to 2012. To achieve this service value, farmers had to use the “biodiverse permanent pasture technique, rich in legumes”. This system involved the no-till seeding of rainfed pastures with a biodiverse mixture of legumes and nonlegumes (up to twenty different species and cultivars, including species with high-self-reseeding capacity, and species able to establish deep roots to deal with cattle pressure and long dry summers), followed by extensive management of stocking rates [40,50]. In eighty Portuguese farms (Figure 2), the rate of SOM increment under this pasture management for a 5-year period was estimated as 0.2% yr−1, corresponding to 5 t CO2eq ha−1 yr−1 [51].
The dairy pasture soils in Australia were very high in SOC concentration (3–7%) in the top 10 cm layer compared with more extensive grazing pastures. The following information from an article entitled ‘Portugal gives green light to pasture carbon farming as a recognized offset’ (Watson, 2010), cited by McKenzie and Watson [52], shows the relevance of the Portuguese technique to managers of degraded land in Australia, where pasture production could provide carbon credits for landholders.
4. Contribution of Belowground Plant Tissue to Carbon (C) Stock in Agroforestry Pasture Soils
It is commonly accepted that an important determinant of plant and ecosystem response to elevated CO2 concentration is plant nutrient status. The C3 plants are more responsive to the increase in atmospheric CO2 than C4 plants. Legumes are C3 plants whereas most grasses are C4. However, C4 grasses in tropical and temperate pastures can sequester a significant amount of C [1]. McSherry and Ritchie [1] explained that C4 grasses may yield more SOC than C3 grasses because of the higher root-to-shoot ratio and greater transfer of photosynthates belowground, greater root density and turnover, and/or possibly higher root exudates. Knowledge on root characteristics that influence nutrient uptake and their response to high CO2 concentration is critical in accurately predicting the long-term plant and ecosystem response to CO2 enrichment [53,54,55].
The tree species in the agroforestry system and the way they are combined influence both the quantity and quality of the biomass returned to the soil [3,4,6]. The contribution of belowground plant biomass to soil C pools (labile or stable) is attracting considerable interest in the particular context of GHG mitigation, but belowground biomass studies in agroforestry systems are problematic, where the dynamics of growth, decay, and turnover of roots is one of the least understood aspects [54]. Therefore, information on C stocks of belowground vegetation components are not usually reported, and those that have been reported lack scientific rigor.
In pasture crops, up to 50% of plant-assimilated C is allocated to belowground tissues [22,56]. The temperate pastures can allocate 24–87% of net primary productivity (NPP) to belowground tissues [57,58]. The major loss of root C via respiration (20–40% of that imported) is associated with plant growth and ion uptake, but fluxes to symbionts are also appreciable, perhaps 10–27% to mycorrhizal fungi [22,35,56,59]. This shows the great potential of roots and mycorrhizal hyphae in SOM and C sequestration. When roots die and decompose, some of their C is released to the atmosphere as CO2 after mineralization, and some remains in soil as SOM. One of the actual challenges is reducing the emission of CO2 from roots and increasing recalcitrant root C to remain in the stable SOM pool.
Storage of SOC is lower in soils of warm and arid climates such as the Mediterranean region relative to those of cool and humid climates, due to the high air temperature and low rainfall. Little SOM decomposition takes place below 10 °C, whereas the optimum temperature for organic material decomposition is 25–30 °C [52,60]. Fine-root decomposition is more sensitive to temperature than leaf litter. Mass loss significantly increases with temperature increase, with an average Q10 of 4.2 estimated for very fine roots (<0.5 mm) compared to 2.36 for leaf litter [59]. However, according to Bonanomi et al. [61], both lignin and (low) N concentration limit decomposition more intensively at low (e.g., 4 °C) than at high (e.g., 24 °C) temperatures. It was hypothesized that the required enzymes to decompose lignin are less efficient at low temperatures. According to Norby and Jackson [62], the turnover rate of fine roots in pastures increased exponentially with temperature, but no relationship was found with rainfall. In Gibraltar in Southern Europe, Gabarrón-Galeote et al. [8] compared three sites for C stocks after land abandonment. Under wetter conditions, the SOC gain in the fine fraction (<50/53 µm) was 59.8% in the fine fraction and in the intermediate fraction (50–250 µm) was 19.4%; in semiarid regions, the SOC gain was 38.2% and 33.3% in the fine and intermediate fractions, respectively. This clearly shows the effect of rainfall on C sequestration.
SOC stock by aboveground temporary spontaneous cover crops in the olive agroforestry in the Andalusia region (Spain) could reach 16 t ha−1 over 10–30 years, with an annual fixation of about 1.5 t SOC ha−1 yr−1 [63]. This shows that areas planted with olives using the same strategy would contribute to the commitment of CO2 reduction. In France, Cardinael et al. [4] estimated a C sequestration rate of 0.24 t C ha−1 yr−1 in an agroforestry system with about 17.8 years. Especially in the Mediterranean region, the C stocks are highly affected by climate change [8]. These authors [8] found that the most stable SOM fraction and the intermediate fraction are the most affected by the predicted increase in temperature and decline in rainfall. On the other hand, the OM in the coarse fraction (>250 µm), mainly as POM, was not affected by the rainfall. These facts indicate that only SOC associated with mineral particles is affected by rainfall, whereas POM is only influenced by land use changes. Recently, Sil et al. [64] observed LULUC changes in the mountain area in the district of Bragança in Northeastern Portugal (lat. 41.9893° to 41.7691°, long. −6.5747° to −6.82292°), with an average annual temperature of 8.5 °C and annual precipitation above 1200 mm, with a positive effect on C storage and sequestration, and an increase in total C storage by 49.2%, at a rate of 1.5 t C ha−1 yr−1. This was mostly due to the expansion of forest areas in the agroforestry systems to the low-SOC-density soils such as the agricultural soils. In improved pastures under the Portuguese agroforestry systems, with an average daily temperature of 15.5–16.8 °C and annual precipitation of 200–750 mm, Teixeira et al. [51] observed a relevant SOM increase after five years’ land use change (2001 to 2005) in all studied cases (80), whereas native pastures did not show a consistent SOM increase (Figure 2).
Root C inputs can be substantial, although the amount declines sharply with soil depth due to the reduced root biomass. Depending on rooting depth, a considerable amount of organic C can be stored below the plow layer where is, therefore, better protection from disturbance, leading to a larger residence time in the soil. Such deep organic C accumulation may be relevant with some trees in agroforestry systems having rooting depths greater than 60 cm.
The decomposition of plant litter is a critical process in global C cycling as it may affect ecosystem productivity and controls the amounts and rates of C sequestered in the ecosystem [58]. Nevertheless, little attention has been paid to root decomposition and element release, and scarce studies are available for Mediterranean conditions [39,42,48,61,65]. It is generally thought that fine roots (<2.0 mm in diameter) of both trees and pasture crops have a relatively fast turnover (days to weeks), and the lignified coarse roots and the larger and thicker hyphae of mycorrhizal fungi usually decompose much more slowly and may thus contribute substantially to C stocks [3,61]. Bonanomi et al. [61] reported that fine roots (<2 mm) only accounted for 14–27% of NPP for an ecosystem C pool because of their rapid turnover. However, Sun et al. [58] and Bonanomi et al. [61] suggested that the turnover of very fine roots (<0.5 mm) of trees (e.g., pine trees) was slower than thicker roots (0.5–2.0 mm) and aboveground litter of the same plant species.
Sun et al. [58], Bonanomi et al. [61], Cao et al. [66], and Wang et al. [67] found strong correlations of fine roots’ decomposition rates with roots’ stoichiometry, but roots’ C quality controlled mostly their decomposition rate. For instance, potential decomposition rates of fine roots (<2.0 mm) were positively related to the concentration of soluble compounds, although negatively related to cellulose concentration in various Mediterranean herbaceous species. According to Fun and Guo [68], fine tree roots, i.e., the lower-order (first- and second-order) roots are expected to decompose more rapidly than higher-order woody roots. However, this prediction has not been tested and may not be correct. Current evidence suggests that lower-order roots may decompose more slowly than higher-order roots in tree species associated with ectomycorrhizal fungi because they are preferentially colonized by fungi and encased by a fungal sheath rich in chitin (a recalcitrant compound) [68]. Sun et al. [58] observed that lower-order roots typically have a lower C/N ratio, particularly in tree species. However, they [58] verified that these roots decomposed more slowly than higher-order roots in both arbuscular mycorrhizal and ectomycorrhizal tree roots in a poor soil, low in nitrogen (N) concentration. They found that the total nonstructural carbohydrate concentration (sugars and starch) of very fine roots (< 0.5 mm) disappeared quickly during the first (82) days of decomposition, followed by the disappearance of acid-hydrolyzable carbohydrate fraction (cellulose and hemicellulose), and finally by the acid-unhydrolyzable carbohydrate fraction (lignin, cutin, suberin, and other aliphatic compounds). Additionally, in nutrient-poor soil, Sun et al. [58] and Bonanomi et al. [61] observed that the very fine roots (<0.5 mm) decomposed slower than the thicker roots (0.5–2.0 mm) in the early and later stages of decomposition. Sun et al. [58] explained the process by a higher concentration of acid-unhydrolyzable carbohydrate fraction, about 15% higher than the 0.5–2.0 mm tree roots, and their lower initial N concentration. Additionally, Bonanomi et al. [61] found a good correlation of decomposition rates and acid-unhydrolyzable carbohydrate concentrations. Both Sun et al. [58] and Bonanomi et al. [61] concluded that very fine roots (<0.5 mm) decomposing in nutrient-poor soils may exacerbate microbial N starvation which is quickly depleted during decomposition. In fact, Carranca et al. [60] found that immobilization of N occurs at an early stage of decomposition, especially in poor soil N where microorganisms use soil N after depletion of tissue N. Bonanomi et al. [61] concluded that a faster decomposition rate occurs in fine roots rich in easily accessible energy-C compounds, and poor in inhibiting compounds. Sun et al. [58], Bonanomi et al. [61], and Fun and Guo [68] concluded that for the quality of C in root litter rather than the initial root litter, composition (stoichiometry) controls most of its decomposition process/rate. Further studies with more plant species, in the field condition and in the long-term, is necessary to validate these findings and clearly explain the mechanisms involved. Although the IPCC tier 1 level calculation [69] considers SOC stock changes to a reference soil depth of 30 cm, from the above discussion, it has been demonstrated that deeper layers should be considered.
Arbuscular mycorrhizal fungi (AMF) are an important group of symbiotic microorganisms associated with various plant species (80% of cultivated plants), comprising 5–50% of the total microbial biomass in soils [59]. This symbiosis is ubiquitous, although with a certain specificity, allowing a better performance and health for most plants, particularly under biotic and abiotic stresses such as drought, high temperature, salinity, depleted nutrients, and the presence of contaminants including heavy metals [70]. Mycorrhizal plants can capture more CO2 from the atmosphere through the photosynthetic process, producing higher levels of photoassimilates that are exuded by roots to the soil, enriching the mycorrhizosphere and contributing to atmospheric C sequestration in soil (mitigation). Hyphal morphology (i.e., wall thickness, width, branching patterns, and turnover), function (i.e., nutrient absorption, plant protection, and soil aggregate formation and stabilization), and longevity vary greatly across and within native and agricultural systems due to changes in the mycorrhizosphere [59]. In studying differences in the architecture and wall thickness of extraradical hyphae, Nichols [59] recognized two types or morphologies: (i) the larger and thicker, more melanized hyphae which are part of the “permanent” fungal network and act as “conduit” or “runner” hyphae, and (ii) the thin-walled, very fine, ephemeral hyphae which are the “absorptive” hyphae and will spread into nutrient-rich microsites in the soil. This author [59] hypothesized that the “absortpive” hyphae would decompose in days or weeks, while the turnover of “runner” hyphae would take years and will contribute to the C sequestration in soil. As Sun et al. [58] and Bonanomi et al. [61] demonstrated, the opposite is expected to happen for very fine roots (<0.5 mm) in trees associated with ectomycorrhizal fungi, especially in a nutrient (N)-depleted soil. The type of mycorrhizal association, i.e., arbuscular, ectomycorrhizal, or ericoid, affects root decomposition, but its effects are not yet fully explored, with particular attention to AMF in pasture species.
Climate change (higher atmospheric CO2 concentration and temperature) will produce larger plants with larger root systems, as a consequence of their size, but with little or no change in C partitioning to them (Table 2). These larger plants may have a higher demand for water and nutrients and, therefore, may deplete the soil in these resources. This can happen in agroforestry systems under the Mediterranean condition. Therefore, there will be more growth of (mycorrhized) roots relative to shoots. If soils become low in nutrients, there may be more exudation from roots and a greater C transfer to mycorrhiza and soil. Changes in mycorrhizal functioning under climate change will have implications for the C cycle, in particular, the amount of C transferred to and stored in the soil. Mycorrhizal fungi will infect roots of C4 grasses more than the C3 grasses in pastures [1]. Differences in mycorrhizal association between C3 and C4 grasses may help to explain SOC differences. In permanent improved pastures in Mediterranean-type areas, the strong competition of grasses and other nonleguminous plants for soil water and nutrients may cause a great exudation and transfer of C into the soil, revealing the great potential of low-input pastures for C sequestration in soil, especially in the presence of higher atmospheric CO2 concentration from climate change. The stronger effect of improved pastures on SOC accumulation beneath trees (e.g., the evergreen oak woodlands in southern Portugal) than in the open field as reported by several authors [42,43,46] may be a result of this hypothesis. However, this remains as an understudied field of research.
In pastures, soil management and environmental conditions influence the diversity and abundance of AMF both in soils and roots. Factors affecting it include the timing, intensity, and duration of plant shoot removal (e.g., grazing) in conjunction with seasonal variation in mycorrhizal formation, the capacity of a host plant to provide the symbiont with photosynthetic products, and the dependence of the host on AMF endophytes [71]. AMF are quite sensitive to soil disturbance and their diversity tends to increase when the intensity of agricultural inputs decreases. Grazing can stimulate the production of mycorrhizal hyphae with a higher excretion of a substance called glomalin into the soil.
An important fraction of the mycorrhizal hyphae mass in pastures is the recalcitrant compound (glomalin) formed by chitin, which can account for the stable SOM pool, lasting for years in the soil [62]. Glomalin is a reddish-brown mycorrhizal fungal hydrophobic glycoprotein, insoluble in water and salt solutions commonly used in soil extractions, and very stable (quite recalcitrant, containing 30–40% organic C). It is linked with soil C storage via its effect on soil aggregate stabilization (the glomalin concentration in soil is highly linearly correlated with soil aggregate water stability) and effectively represents an important soil C pool [72,73]. As Six et al. [34] indicated, mycorrhizal (and saprophytic) fungi are the most important soil microorganisms involved in the formation and stabilization of aggregates [34]. Glomalin is deposited outside the hyphal walls of the extraradical mycelium and on adjacent soil particles, and appear to act as a long-term soil cement agent contributing to forming microaggregates [72,73,74]. Plant species differ from each other in influencing aggregate formation, but many factors are responsible for this effect (namely the root size and colonization, plant nutrition, soil structure, and climatic conditions) [75]. Soil aggregation is very fast through AMF root colonization. Marcel et al. [75] verified that microcosms inoculated with AMF in agricultural soils formed 3.2% more aggregates (>1 mm) than microcosms without AMF. In order to use the glycoprotein concentration as a specific assessment of soil quality and management impact on soil C sequestration, it is necessary to relate the concentration of this molecule to the increase in aggregate stability over time. Rillig et al. [73] observed that the percentage of large macroaggregates (>2 mm) increased significantly from 10% in agricultural soil to 30% in afforested soil, showing a strong positive response of glomalin to the land use change. Therefore, glomalin may be used as a useful sensitive indicator of soil C changes in response to land use practices, but further studies are needed to demonstrate these findings [73].
Standing stocks of AMF hyphae in soil were estimated of the order of 0.05–0.90 t C ha−1, where glomalin apparently contributed to 0.4–6% of this biomass [52,59]. Together with the glomalin in mycorrhizal exudates, when the hyphae senesce glomalin in the walls is also deposited in the soil where it accumulates. Rillig et al. [73] estimated the total contribution of glomalin C to the total SOC pool to vary from 3.8% to 7.8%, depending on land use type and total SOC (from agricultural to afforested and natural soils). They observed that about one half of the glomalin was still detectable after 400 days of incubation at 25 °C and 60% water-holding capacity, suggesting that 50% of this protein was in the labile fraction of the soil C pool and was lost in the first phase of decomposition; the remaining part was of the recalcitrant fraction. By correcting data for temperature in the field, a mean residence time of 2.67 times slower was reported, suggesting that glomalin can be much longer-lived in the field than expected for plant-derived proteins. In a study with a tropical forest soil and using the 14C data, Rillig et al. [76] reported a turnover of 6–42 years for glomalin.
5. New Approaches for Appropriate Increase in Carbon (C) Sequestration in Mediterranean Agroforestry Soils
To maintain agricultural ecosystem functions with mitigation of atmospheric CO2 and a consistent supply of food, feed, fiber, and energy to a growing global population, appropriate land use and management systems need to be developed and implemented in the Mediterranean agroforestry systems. No-tillage practice along with permanent improved pasture cultivation increases the percentage of water-stable aggregates within the soil by increasing mycorrhizal hyphal length and microbial exudates in the mycorrhizosphere. Higher levels of C sequestration in no-tilled soils are possible, since not only C is allocated belowground to hyphal networks with the formation of highly stable glomalin, but also because the SOM occluded within aggregates appears to have a greater turnover time than the free SOM.
Nevertheless, particular challenges remain in demonstrating the relevance of mycorrhizal (i.e., glomalin) contribution for mitigating the atmospheric CO2 in permanent pastures. Since mycorrhizal groups differ in their growth rate and recalcitrant chitin content, these shifts can have implications on C storage in pasture soils, and more research on mycorrhizal physiology is needed before any prediction can be made. Although Rillig et al. [73] found a decreasing total glomalin concentration from horizon A to C in agricultural, afforested, and natural soils, still, there is a lack of knowledge on glomalin distribution in agroforestry soil profiles, since most (few) studies focused on the top layers (0–20 cm), where the largest C pools are detected (mostly as labile SOM).
Information is lacking on stocks of organic C in deeper layers where most tree roots occur and where supply of C substantially is generated through root exudates and fine-root turnover. Assessing soil C for global change scenarios requires an understanding of C pools in deeper layers, especially for both native and improved pastures, as is the case for Mediterranean agroforestry systems. The interaction of pastures with trees in agroforestry systems also needs further studies with respect to the contribution of mycorrhizal systems to the efficiency of soil C storage. Increasing pasture productivity and root mass inputs to the soil is one key to enhance C stock, for example through the application of phosphorus and the sowing of productive pasture crops, possibly in combination with bred plant legumes, as in the “biodiverse Portuguese technique”. All this information is of utmost importance for policymakers and managers to establish guidelines regarding pastures with tree management in agroforestry systems towards better ecosystem stability and an improvement in service value that farmers and society have to take into account.
Author Contributions
C.C. contributed to the article’s conception and manuscript preparation; F.P. contributed to manuscript preparation and revision; M.M. contributed substantially to the article’s preparation and revision. All authors approved the submitted version and agreed to be personally accountable for the author’s own contributions and for questions related to the accuracy or integrity of any part of the work, even ones in which the author was not personally involved and documented in the literature. All authors have read and agreed to the published version of the manuscript.
Funding
This review was funded by the Ref. 4551-GREEN-IT “Bioresources for Sustainability” project to cover publication costs.
Conflicts of Interest
The authors declare no conflict of interest.
References
- McSherry, M.; Ritchie, M.E. Effects of grazing on grassland soil carbon: A global review. Glob. Chang. Biol. 2013, 19, 1347–1357. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC). Contribution of Working Group II (WGII): Climate Change 2007: Impacts, Adaptation and Vulnerability; Parry, M., Canziani, O., Palutikof, J., van der Linden, P., Hanson, C., Eds.; Published for the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Nair, P.K.R.; Kumar, B.M.; Nair, V.D. Agroforestry as a strategy for carbon sequestration. J. Plant Nutr. Soil. Sci. 2009, 172, 10–23. [Google Scholar] [CrossRef]
- Cardinael, R.; Umulisa, V.; Toudert, A.; Olivier, A.; Bockel, L.; Bernoux, M. Revisiting IPCC Tier 1 coefficients for soil organic and biomass carbon storage in agroforestry systems. Environ. Res. Lett. 2018, 13, 124020. [Google Scholar] [CrossRef]
- World Meteorological Organization (WMO); United Nations Environment Programme; Intergovernmental Panel on Climate Change. Emissions Scenarios; IPCC Special Report; IPCC: Geneva, Switzerland, 2000; ISBN 92-9169-113-5. [Google Scholar]
- Nair, P.K.R. Carbon sequestration studies in agroforestry systems: A reality—Check. Agrofor. Syst. 2012, 86, 243–253. [Google Scholar] [CrossRef]
- Feliciano, D.; Ledo, A.; Hiller, J.; Nayak, D.R. Which agroforestry options give the greatest soil and above ground carbon benefits in different world regions? Agric. Ecosyst. Environ. 2018, 254, 117–129. [Google Scholar] [CrossRef]
- Gabarrón-Galeote, M.A.; Trigalet, S.; van Wesemael, B. Effect of land abandonment on soil organic carbon fractions along a Mediterranean precipitation gradient. Geoderma 2015, 249–250, 69–78. [Google Scholar] [CrossRef]
- Chiti, T.; Gardin, L.; Perugini, L.; Quaratino, R.; Vaccari, F.P.; Miglietta, F.; Valentini, R. Soil organic carbon stock assessment for the different cropland land uses in Italy. Biol. Fertil. Soils 2011, 48, 9–17. [Google Scholar] [CrossRef]
- Six, J.; Conant, R.T.; Paul, E.A.; Paustian, K. Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils. Plant Soil 2002, 241, 155–176. [Google Scholar] [CrossRef]
- Dungait, J.A.J.; Hopkins, D.W.; Gregory, A.S.; Whitmore, A.P. Soil organic matter turnover is governed by accessibility not recalcitrance. Glob. Chang. Biol. 2012, 18, 1781–1796. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). The Regional Impacts of Climate Change: An Assessment of Vulnerability; Watson, R.T., Zinyowera, M.C., Moss, R.H., Eds.; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Franzluebbers, A.J.; Arshad, M.A. Particulate Organic Carbon Content and Potential Mineralization as Affected by Tillage and Texture. Soil Sci. Soc. Am. J. 1997, 61, 1382–1386. [Google Scholar] [CrossRef]
- Huang, Z.; Davis, M.R.; Condron, L.M.; Clinton, P. Soil carbon pools, plant biomarkers and mean carbon residence time after afforestation of grassland with three tree species. Soil Biol. Biochem. 2011, 43, 1341–1349. [Google Scholar] [CrossRef]
- Bayer, C.; Mielniczuk, J.; Martin-Neto, L.; Ernani, P.R. Stocks and humification degree of organic matter fractions as affected by no-tillage on a subtropical soil. Plant Soil 2002, 238, 133–140. [Google Scholar] [CrossRef]
- Cambardella, C.A.; Elliott, E.T. Particulate Soil Organic-Matter Changes across a Grassland Cultivation Sequence. Soil Sci. Soc. Am. J. 1992, 56, 777–783. [Google Scholar] [CrossRef]
- Batjes, N.H. Management Options to Reduce CO2—Concentrations in the Atmosphere by Increasing Carbon Sequestration in the Soil; NRS Report no. 410200031; International Soil Reference and Information Centre (ISRIC): Wageningen, The Netherlands, 1999. [Google Scholar]
- Cheng, H.H.; Kimble, J.M. Characterization of soil organic carbon pools. In Assessment Methods for Soil Carbon; Lal, R., Kimble, J.M., Follett, R.F., Stewart, B.A., Eds.; Lewis Publishers: London, UK, 2001; pp. 117–129. [Google Scholar]
- Pedra, F.; Plaza, C.; Fernández, J.M.; García-Gil, J.C.; Polo, A. Effects of municipal solid waste compost and sewage sludge on chemical and spectroscopic properties of humic acids from a sandy Haplic Podzol and a clay loam Calcic Vertisol in Portugal. Waste Manag. 2008, 28, 2183–2191. [Google Scholar] [CrossRef]
- Lehmann, J.; Kleber, M. The contentious nature of soil organic matter. Nature 2015, 528, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Ponge, J.-F. Humus: Dark side of life or intractable “aether”? Pedosphere 2022, 32, 660–664. [Google Scholar] [CrossRef]
- Govaerts, B.; Verhust, N.; Castellanos-Navarrete, A.; Sayre, K.D.; Dixon, J. Conservation agriculture and soil carbon sequestration: Between myth and farmer reality. Crit. Rev. Plant Sci. 2009, 28, 97–122. [Google Scholar] [CrossRef]
- McFariane, K. Belowground Carbon Storage and Soil Organic Matter Quality Following Fertilizer and Herbicide Applications in Ponderosa Pine Plantations Along a Site-quality Gradient in Northern California; Oregon State University: Corvallis, OR, USA, 2007. [Google Scholar]
- Wander, M.M.; Bidart, M.G.; Aref, S. Tillage Impacts on Depth Distribution of Total and Particulate Organic Matter in Three Illinois Soils. Soil Sci. Soc. Am. J. 1998, 62, 1704–1711. [Google Scholar] [CrossRef]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Yang, X.M.; Xie, H.; Drury, C.F.; Reynolds, W.D.; Yang, J.Y.; Zhang, X.D. Determination of organic carbon and nitrogen in particulate organic matter and particle size fractions of Brookston clay loam soil using infrared spectroscopy. Eur. J. Soil Sci. 2012, 63, 177–188. [Google Scholar] [CrossRef]
- Cotrufo, M.F.; Soong, J.L.; Horton, A.J.; Campbell, E.E.; Haddix, M.L.; Wall, D.H.; Parton, W.J. Formation of soil organic matter via biochemical and physical pathways of litter mass loss. Nat. Geosci. 2015, 8, 776–779. [Google Scholar] [CrossRef]
- Santoiemma, G. Recent methodologies for studying the soil organic matter. Appl. Soil Ecol. 2018, 123, 546–550. [Google Scholar] [CrossRef]
- Khanna, P.K.; Ludwig, B.; Bauhus, J.; O’Hara, C. Assessment and significance of labile organic C pools in forest soils. In Assessment Methods for Soil Carbon; Lal, R., Kimble, J.M., Follett, R.F., Stewart, B.A., Eds.; Lewis Publishers: London, UK, 2001; pp. 167–182. [Google Scholar]
- Hassink, J.; Whitmore, A.P. A model of the physical protection of organic matter in soils. Soil Sci. Soc. Am. J. 1997, 61, 131–139. [Google Scholar] [CrossRef]
- Neff, J.C.; Townsend, A.R.; Gleixner, G.; Lehman, S.; Turnbull, J.J.; Bowman, W.D. Variable effects of nitrogen additions on the stability and turnover of organic carbon. Nature 2002, 419, 915–917. [Google Scholar] [CrossRef] [PubMed]
- Covaleda, S.; Gallardo, F.; García-Oliva, F.; Kirchmann, H.; Prat, C.; Bravo, M.; Etchevers, J.D. Land-use effects on the distribution of soil organic carbon within particle-size fractions of volcanic soils in the Transmexican Volcanic Belt (Mexico). Soil Use Manag. 2011, 27, 186–194. [Google Scholar] [CrossRef]
- Cerdà, A. Soil aggregate stability under different Mediterranean vegetation types. Catena 1998, 32, 73–86. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil & Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Lavalle, P.; Spain, A.; Fonte, S.; Bedano, J.C.; Blanchart, E.; Galindo, V.; Grimaldi, M.; Jimenez, J.J.; Velasquez, E.; Zangerlé, A. Soil aggregation, ecosystem engineers and the C cycle. Acta Oecol. 2020, 105, 103561. [Google Scholar] [CrossRef]
- Costa, A.; Madeira, M.; Santos, J.L.; Oliveira, Â. Change and dynamics in Mediterranean evergreen oak woodlands landscapes of Southwestern Iberian Peninsula. Landsc. Urban Plan. 2011, 102, 164–176. [Google Scholar] [CrossRef]
- Nunes, M.R.; Karlen, D.L.; Veum, K.S.; Moorman, T.B. A SMAF assessment of U.S. tillage and crop management strategies. Environ. Sustain. Indic. 2020, 8, 100072. [Google Scholar] [CrossRef]
- Hernández-Esteban, A.; López-Díaz, M.L.; Cáceres, Y.; Moreno, G. Are sown legume-rich pastures effective allies for the profitability and sustainability of Mediterranean dehesas? Agrofor. Syst. 2018, 93, 2047–2065. [Google Scholar] [CrossRef]
- Simón, N.; Montes, F.; Díaz-Pinés, E.; Benavides, R.; Roig, S.; Rubio, A. Spatial distribution of the soil organic carbon pool in a Holm oak dehesa in Spain. Plant Soil 2012, 366, 537–549. [Google Scholar] [CrossRef] [Green Version]
- Crespo, D.G. The Role of Pasture Improvement in the Rehabilitation of the "Montado/Dehesa" System and in Developing Its Traditional Products. In Animal Products from the Mediterranean Area; Ramalho Ribeiro, J.M.C., Horta, A.E.M., Mosconi, C., Rosati, A., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2006; pp. 185–197. [Google Scholar]
- Rice, C.W. Management effects on biological and physical controls of soil C sequestration. In Proceedings of the 4th International Congress of the European Soil Science Societies Eurosoil 2012: Soil Science for the Benefit of Mankind and Environment, Bari, Italy, 2–6 July 2012. [Google Scholar]
- Gómez-Rey, M.X.; Garcês, A.; Coutinho, J.; Madeira, M. Soil organic C accumulation and N availability under improved pastures established in Mediterranean oak woodlands. Soil Use Manag. 2012, 28, 497–507. [Google Scholar] [CrossRef]
- Gómez-Rey, M.X.; Gonzales-Prieto, S.J.; Madeira, M.; Coutinho, J. Soil C and N dynamics in Mediterranean oak woodland with schrub encroachment. Plant Soil 2013, 371, 339–354. [Google Scholar] [CrossRef]
- Escudero, A.; Garcia, B.; Gomez, J.M.; Luis, E. The nutrient cycling in Quercus rotundifolia and Quercus pyrenaica ecosystems (“dehesa”) of Spain. Acta Oecol./Oecol. Plant. 1985, 6, 73–86. [Google Scholar]
- Moreno, G.; Obrador, J.J.; Cubera, E.; Dupraz, C. Fine root distribution in dehesas of Central–Western Spain. Plant Soil 2005, 277, 153–162. [Google Scholar] [CrossRef]
- Pulido-Fernández, M.; Schnabe, S.; Lavado-Contador, J.F.; Mellado, I.M.; Pérez, R.O. Soil organic matter of Iberian open woodland rangelands as influenced by vegetation cover and land management. Catena 2013, 109, 13–24. [Google Scholar] [CrossRef]
- Howlett, D.S.; Moreno, G.; Mosquera Losada, M.R.; Nair, P.K.R.; Nair, V.D. Soil carbon storage as influenced by tree cover in the Dehesa cork oak silvopature of central–western Spain. J. Environ. Monit. 2011, 13, 1897–1904. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, A.R.; Costa e Silva, F.; Correia, A.C.; Bicho, M.C.; Madeira, M.; Coutinho, J. Do improved pastures enhance soil quality of cork oak woodlands in the Alentejo region (Portugal)? Agrofor. Syst. 2020, 94, 125–136. [Google Scholar] [CrossRef]
- Teixeira, R.F.M.; Proença, V.; Crespo, D.; Domingos, T. A conceptual framework for the analysis of engineered biodiverse pastures. Ecol. Eng. 2015, 44, 187–196. [Google Scholar] [CrossRef]
- Hernandez–Esteban, A.; Moreno, M.G. Establishing pastures rich in legumes. Agroforesty Innov. 2017, 1, 2. Available online: https://www.agforward.eu (accessed on 1 September 2022).
- Teixeira, R.F.M.; Domingos, T.; Costa, A.P.S.V.; Oliveira, R.; Farropas, L.; Calouro, F.; Barradas, A.M.; Carneiro, J.P.B.G. Soil organic matter dynamics in Portuguese natural and sown rainfed grasslands. Ecol. Model. 2011, 222, 993–1001. [Google Scholar] [CrossRef]
- McKenzie, D.; Watson, W. Soil Carbon Sequestration under Pasture in Australian Dairy Regions. In Dairy Australia; Project MCK13538; McKenzie Soil Management Pty. Ltd.: Orange, NSW, Australia, 2010; Volume 42. [Google Scholar]
- BassiriRad, H. Kinetics of nutrient uptake by roots: Responses to global climate change. New Phytol. 2000, 147, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Carranca, C. The contribution of Portuguese agriculture to the climate change, mitigation and adaptation strategies for the sector. In Proceedings of the Advances in Climate Changes, Global Warming, Biological Problems and Natural Hazards, 3rd International Conference on Climate Changes, Global Warming, Biological Problems (CGB’10), and 3rd WSEAS International Conference on Natural Hazards (NAHA’10), University of Algarve, Faro, Portugal, 3–5 November 2010; pp. 48–56. [Google Scholar]
- Strand, A.E.; Pritchard, S.G.; McCormack, M.L.; Davis, M.A.; Oren, R. Irreconcilable differences: Fine-root life spans and soil carbon persistence. Science 2008, 319, 456–458. [Google Scholar] [CrossRef]
- Farrar, J.F.; Jones, D.L. The control of carbon acquisition by roots. Research review. New Phytol. 2000, 147, 43–53. [Google Scholar] [CrossRef]
- Gill, R.A.; Jackson, R.B. Global patterns of root turnover for terrestrial ecosystems. New Phytol. 2000, 147, 13–31. [Google Scholar] [CrossRef]
- Sun, T.; Mao, Z.; Han, Y. Slow decomposition of very fine roots and some factors controlling the process: A 4–year experiment in four temperate tree species. Plant Soil 2013, 372, 445–458. [Google Scholar] [CrossRef]
- Nichols, K.A. Indirect contributions of AM fungi and soil aggregation to plant growth and protection. In Mycorrhizae: Sustainable Agriculture and Forestry; Siddiqui, A., Akhtar, M.S., Futai, K., Eds.; Springer Science + Business Media BV: Berlin/Heidelberg, Germany, 2008; pp. 177–194. [Google Scholar]
- Carranca, C.; Rocha, I.; de Varennes, A.; Oliveira, A.; Pampulha, M.E.; Torres, M.O. Effect of tillage and temperature on potential nitrogen mineralization and microbial activity and microbial numbers of lupine amended soil. Agrochimica 2009, LIII, 183–195. [Google Scholar]
- Bonanomi, G.; Idbella, M.; Zotti, M.; Santorufo, L.; Motti, R.; Maisto, G.; De Marco, A. Decomposition and temperature sensitivity of fine root and leaf litter of 43 mediterranean species. Plant Soil 2021, 464, 453–465. [Google Scholar] [CrossRef]
- Norby, R.J.; Jackson, R.B. Root dynamics and global change: Seeking an ecosystem perspective. Research review. New Phytol. 2000, 147, 3–12. [Google Scholar] [CrossRef] [Green Version]
- Torrús–Castillo, M.; Domouso, P.; Herrera–Rodriguez, J.M.; Calero, J. Aboveground carbon fixation and nutrient retention in temporary spontaneous cover crops in olive groves of Andalusia. Front. Environ. Sci. Front. Environ. Sci. 2022, 10, 1–13. [Google Scholar] [CrossRef]
- Sil, A.; Fonseca, F.; Gonçalves, J.; Honrado, J.; Marta-Pedroso, M.; Alonso, J.; Ramos, M.; Azevedo, J.C. Analysing carbon sequestration and storage dynamics in a changing mountain landscape in Portugal: Insights for management and planning. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2017, 82–104. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, C.; Madeira, M. Root decomposition process in Mediterranean conditions. In Proceedings of the Transactions of the 15th World Congress of Soil Science, Acapulco, Mexico, 10–16 July 1994; 4b Commission III: Poster Sessions. International Society of Soil Science/Mexican Society of Soi Science: Acapulco, Mexico, 1994; pp. 102–103. [Google Scholar]
- Cao, Y.; Li, Y.; Zhang, G.; Zhang, J.; Chen, M. Fine root C:N:P stoichiometry and its driving factors across forest ecosystems in northwestern China. Sci. Total Environ. 2020, 737, 140299. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chen, Y.; Zhou, Y.; Zheng, H.; Xu, Z.; Tan, B.; You, C.; Zhang, L.; Li, H.; Guo, L.; et al. Litter chemical traits strongly drove the carbon fractions loss during decomposition across an alpine treeline ecotone. Sci. Total Environ. 2020, 753, 142287. [Google Scholar] [CrossRef]
- Fan, P.; Guo, D. Slow decomposition of lower order roots: A key mechanism of root carbon and nutrient retention in the soil. Oecologia 2010, 163, 509–515. [Google Scholar] [CrossRef]
- Intergovernmental Panel on Climate Change (IPCC). IPCC Guidelines for National Greenhouse Gas Inventories. Report 2006; Eggelston, S., Buendia, L., Miwa, K., Ngara, T., Tanabe, K., Eds.; Institute for Global Environmental Strategies (IGES) for the IPCC: Hayama, Japan, 2006; Volume 4, ISBN 4-88788-032-4. [Google Scholar]
- Cruz, C.; Carranca, C. A colonização micorrízica no uso eficiente do azoto pelas culturas hortícolas. Rev. APH 2010, 9–12. [Google Scholar]
- Benthlefalvay, G.J. Review article. Mycorrhizae in the agricultural plant-soil system. Symbiosis 1992, 14, 413–425. [Google Scholar]
- Wright, S.F.; Starr, J.L.; Paltineanu, E.C. Changes in aggregate stability and concentration of glomalin during tillage management transition. Soil Sci. Soc. Am. J. 1999, 63, 1825–1829. [Google Scholar] [CrossRef]
- Rillig, M.C.; Ramsey, P.W.; Morris, S.; Paul, E.A. Glomalin, an arbuscular-mycorrhizal fungal soil protein, responds to land-use change. Plant Soil 2003, 253, 293–299. [Google Scholar] [CrossRef]
- Wang, P.; Liu, J.-H.; Xia, R.-X.; Wu, Q.-S.; Wang, M.-Y. Arbuscular mycorrhizal development, glomalin-related soil protein (GRSP) content, and rhizosphere phosphatase activity in citrus orchards under different types of soil management. J. Plant Nutr. Soil Sci. 2011, 174, 65–72. [Google Scholar] [CrossRef]
- Marcel, G.A.H.; Streitwolf-Engel, R.; Riedl, R.; Siegrist, S.; Neudecker, A.; Ineichen, K.; Boller, T.; Wiemken, A.; Sanders, I.R. The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland. New Phytol. 2006, 172, 739–752. [Google Scholar] [CrossRef]
- Rillig, M.C.; Wright, S.F.; Nichols, K.A.; Schmidt, W.F.; Torn, M.S. Large contribution of arbuscular mycorrhizal fungi to soil carbon pool in tropical soils. Plant Soil 2001, 233, 167–177. [Google Scholar] [CrossRef]
Figure 1.
Partial view of a Portuguese agroforestry system with Quercus sp. and a biodiverse pasture.
Figure 1.
Partial view of a Portuguese agroforestry system with Quercus sp. and a biodiverse pasture.

Figure 2.
Variation in actual soil organic matter (SOM) at 0–20 cm soil depth, in two different pastures in Portugal, comparing data of 2001 and 2005). (Adapted from [51] Teixeira et al., 2011.)
Figure 2.
Variation in actual soil organic matter (SOM) at 0–20 cm soil depth, in two different pastures in Portugal, comparing data of 2001 and 2005). (Adapted from [51] Teixeira et al., 2011.)


{kind=link}
{kind=link}
Table 1.
Ranges in the amount and turnover times of various types of soil organic matter (SOM) stored in agricultural soils (Source: Batjes [17]).
Table 1.
Ranges in the amount and turnover times of various types of soil organic matter (SOM) stored in agricultural soils (Source: Batjes [17]).
| Type of SOM | Proportion of Total SOM (%) | Turnover Time (Year) | C-Pool |
|---|---|---|---|
| Microbial biomass | 2–5 | 0.1–0.4 | labile |
| Litter | - | 1–3 | rapid |
| Particulate organic matter (POM) | 18–40 | 5–20 | moderate |
| Light fraction | 10–30 | 1–15 | moderate |
| Inter-micro-aggregate a | 20–35 | 5–50 | moderate to slow |
| Intra-micro-aggregate: b | |||
| Physically sequestered | 20–40 | 50–1000 | passive |
| Chemically sequestered | 20–40 | 1000–3000 | passive |
a within macroaggregates, but external to microaggregates, including POM, light fraction, and microbial carbon (C); b within microaggregates, including sequestered light fraction and microbial derived C.
Table 2.
Growth and root metabolism of barley at 14 days after germination under two CO2 concentrations (Source: Farrar and Jones [56]).
Table 2.
Growth and root metabolism of barley at 14 days after germination under two CO2 concentrations (Source: Farrar and Jones [56]).
| Variables | CO2 Concentration (ppm) | |
|---|---|---|
| 350 | 700 | |
| Dry weight (mg) | 154 | 222 |
| Root weight ratio | 0.31 | 0.31 |
| Rate of elongation of seminal axis (cm h−1) | 0.12 | 0.18 |
| Number of nodal roots | 3.4 | 5.8 |
| Rate of lateral root production (h−1) | 1.1 | 1.8 |
| Carbohydrate content of seminal axes (mg g−1 FW) | 3.2 | 4.0 |
| Respiration of seminal axes (nmol g−1 s−1) | 2.7 | 3.9 |
| Carbohydrate content of root tips (mg g−1 FW) | 17.3 | 37.9 |
| Respiration of root tips (pmol per tip s−1) | 22 | 33 |
FW = fresh weight.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Carranca, C.; Pedra, F.; Madeira, M. Enhancing Carbon Sequestration in Mediterranean Agroforestry Systems: A Review. Agriculture 2022, 12, 1598. https://doi.org/10.3390/agriculture12101598
AMA Style
Carranca C, Pedra F, Madeira M. Enhancing Carbon Sequestration in Mediterranean Agroforestry Systems: A Review. Agriculture. 2022; 12(10):1598. https://doi.org/10.3390/agriculture12101598
Chicago/Turabian StyleCarranca, Corina, Filipe Pedra, and Manuel Madeira. 2022. "Enhancing Carbon Sequestration in Mediterranean Agroforestry Systems: A Review" Agriculture 12, no. 10: 1598. https://doi.org/10.3390/agriculture12101598
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.